栽培生理
摘要:4-香豆酸辅酶A连接酶(4CL)是植物苯丙烷代谢途径的核心酶,是类黄酮和木质素生物合成的关键酶,对葡萄果实品质和抗逆境胁迫形成有重要作用。通过生物信息学分析,从葡萄中共鉴定出15个4CL基因家族成员(Vv4CL01-Vv4CL15),主要分布于Chr13和Chr18等7条染色体,系统发育分析将其分成4个亚族(I-IV)。顺式作用元件分析发现4CL基因启动子区富集与生长发育、激素和胁迫响应相关的作用元件。共线性分析表明片段复制是家族扩张的主要驱动力。qRT-PCR结果发现Vv4CL02和Vv4CL06在‘巨峰’葡萄的穗轴中高水平表达;Vv4CL03在‘阳光玫瑰’葡萄的幼嫩果粒和茎中表达水平最高。该研究为进一步探究葡萄4CL基因家族参与响应多种逆境胁迫的作用机制提供参考。
关键词:葡萄;Vv4CL基因家族;组织特异性;表达分析
葡萄品质形成高度依赖次生代谢产物的精准调控,果皮中的花色苷决定果实色泽与抗氧化活性,果肉中的类黄酮影响口感与营养品质,而木质素则参与穗轴机械支撑结构构建及果实硬度形成[1]。4-香豆酸辅酶A连接酶(4CL)是植物苯丙烷代谢途径的核心酶,可催化羟基肉桂酸类化合物与辅酶A结合形成辅酶A酯,为木质素、黄酮类等多种次生代谢产物的合成提供关键前体[2]。作为苯丙烷代谢的分支调控节点,4CL通过活化不同底物将代谢流导向特定分支途径:其产物4-香豆酰辅酶A既可参与木质素的生物合成[3],通过氧化聚合形成植物细胞壁的结构成分,增强植物的机械强度和抗病性[4];也可作为黄酮类化合物(如花青素、黄酮醇)的合成起点,影响果实色泽、抗氧化活性及紫外线防御[5];同时,其衍生物还可进一步生成芪类化合物、酚酸类及部分芳香物质,在植物生长发育、抗逆响应和品质形成中发挥重要作用。
4CL基因在1988年首次在欧芹(Petroselinum hortense)被克隆出来,随后4CL在更多物种中得到系统的鉴定和研究[6]。4CL基因通常以多基因家族形式存在,成员间因基因结构、启动子顺式作用元件及系统进化关系的差异而功能分化[7-8]。葡萄等经济作物中,4CL基因家族成员的表达具有组织特异性,如在果皮中高表达的成员可能驱动花色
苷积累,而在木质化组织活跃的成员则促进木质素沉积[9-10]。
目前,拟南芥[11]、水稻[12]、桑树[13]、水芹[14]、甘草[5]和茶树[15]等植物的4CL基因家族已得到系统性研究,揭示了基因家族扩张与功能分化的进化模式。有研究表明拟南芥中At4CL1、At4CL2、At4CL4参与了木质素的合成,At4CL3和水稻中的Os4CL2参与了类黄酮的生物合成[11-12]。苹果Md4CL10和Md4CL23的表达量与其果实着色和花青素积累程度成正比[16]。然而,葡萄4CL基因家族的研究仍较为滞后,现有成果多集中于分子克隆与初步表达分析,缺乏全基因组层面的系统性鉴定和进化关联解析。虽然葡萄基因组资源的发布为基因家族分析提供了基础,但当前研究仍依赖单一参考基因组,可能忽视结构变异(SVs)对基因功能的影响,例如栽培葡萄因驯化导致的抗病基因丢失与次生代谢通路基因的多样性缩减问题[17]。近期,中国科学家通过构建葡萄单倍型超级泛基因组,揭示了欧亚种群与北美种群的频繁杂交事件及东亚种群的未充分利用潜力,为4CL基因的跨物种比较和驯化选择印记分析提供了新方向[18]。然而,葡萄4CL基因在泛基因组框架下的进化-功能关联图谱仍待构建,其在果实品质形成中的调控网络亦未明确。
本研究聚焦葡萄4CL基因家族在苯丙烷代谢途径中的关键作用,通过全基因组鉴定系统性分析葡萄4CL家族成员的基因结构、保守结构域、启动子顺式作用元件及系统发育进化,并检测该基因家族在不同组织器官中的表达模式,为阐明葡萄4CL基因的生物学功能及其调控木质素、黄酮类化合物合成的分子机制提供理论依据,同时为葡萄抗性改良与品质育种提供靶向基因资源。
2. 试验材料与方法
2.1试验材料
试验材料为北辰双街现代农业科技园日光温室栽培的‘阳光玫瑰’与‘巨峰’。2024年5月17日取样,取两个品种距基部第三片叶的腋芽、距基部第二节茎、幼嫩卷须、果粒(2~3 mm)、穗轴;2024年5月23日取样,取根、距顶端第三片叶、果粒(13~18 mm)。取样后立即放入液氮中冷冻,用研钵研磨成果粉后置于-80℃冰箱中保存备用。
2.2 试验方法
2.2.1 4CL基因家族成员的鉴定
从Phytozome数据库(https://phytozome-next.jgi.doe.gov/)下载‘黑比诺’葡萄基因组PN40024 v2.1的FA和gff3文件及拟南芥等参考物种的4CL基因序列。根据拟南芥4CL基因序列,使用在线工具TBtools和NCBI的ProteinBlast,经Blast初步筛选与双向Blast精准鉴定从‘黑比诺’葡萄基因组中筛选出4CL基因家族的候选基因序列和蛋白质序列,再分别使用DTU Health Tech网站(https://services. healthtech.dtu.dk/services/TMHMM-2.0/)、Expasy网站(http://www.expasy.org)和WoLF PSORT网站(https://wolfpsort.hgc.jp/)进行蛋白长度、跨膜结构域预测和等电点/分子量等蛋白理化性质分析,以及亚细胞定位预测的分析。
2.2.2 染色体分布与成员序列保守特征
利用在线工具TBtools将鉴定到的候选基因在染色体上进行可视化,并依据其在染色体上的位置依次命名,并进行基因结构注释分析。在GSDS2.0网站https://gsds.gao-lab.org/index.php分析基因序列的保守结构域,利用MEME网站(https://meme-suite.org/meme/tools/meme)基于motif分析预测其蛋白序列的结构域,在InterProScan网站(https://www.ebi.ac.uk/interpro/),筛选出目的保守结构域,再运用TBtools软件进行可视化。
2.2.3 基因成员的系统进化树
从NCBI数据库下载拟南芥(Arabidopsis thaliana)、葡萄(Vitis vinifera)、蓝莓(Vaccinium darrowi)、香蕉(Musa acuminata)、凤梨(Ananas comosus)的4CL蛋白序列,使用MEGA软件进行多重序列比对,再利用TBtools (Quixk Run TrimAL工具)进行比对结果修剪;将比对结果导入MEGA软件进行邻位相连(Neighbor-Joining)法设置Bootstrap为1000次重复构建系统发育树;最后使用在线网站ITOL(https://itol.embl.de/)进行美化。
2.2.4 基因顺式作用元件分析
使用TBtools软件(GXF Sequences Extract工具)提取葡萄4CL基因家族成员转录起始位点上游1500 bp的序列,提交到PlantCare网站
(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析顺式作用元件,然后在TBtools (Basic Biosequence View工具)可视化。
2.2.5 共线性分析
从NCBI网站上下载拟南芥、葡萄、蓝莓、香蕉、凤梨的基因组文件.fa,利用TBtools (One Step MCScanX-super Fast)插件进行葡萄种内和两两物种间的共线性分析,分别利用TBtools 的Advanced Circos和Multiple Synteny Plot插件进行可视化。
2.2.6 荧光定量PCR检测
采用RNAprepPure多糖多酚植物总RNA提取试剂盒(天根生化科技(北京)有限公司,产品号:DP220706),根据试剂盒说明书提取各组织样品的总RNA。使用FastKing一步法除基因组cDNA第一链合成预混试剂合成第一链cDNA,反转录1 μg的总RNA,反转录体系20 μl。利用在线网站NCBI设计4CL基因家族荧光定量引物(引物序列见表1),以反转录得到的cDNA产物作为模板,使用QuantStudio7 Flex 实时荧光定量PCR系统(Life Iechnologies USA),采用SYBR GreenI染料法进行实时荧光定量PCR。反应程序为95℃ 30 s;95℃ 10 s;60℃ 20 s;72℃ 30 s,40个循环。基因相对表达量按照2-△△Ct方法计算,两个品种均以“距基部第三片叶的腋芽”作为对照组

3. 结果与分析
3.1 葡萄4CL基因家族成员的鉴定
以拟南芥7个4CL基因为参照,从葡萄基因组中鉴定了15个4CL家族成员(Vv4CL01~Vv4CL15)。从表2可知:蛋白长度493~570 aa,分子量53.02~61.20 kDa,Vv4CL09蛋白序列最长、Vv4CL06和07的最短;等电点两极分化,部分为碱性蛋白,部分为酸性蛋白。Vv4CL03、04、05、15蛋白含1~2个跨膜螺旋且拓扑结构有差异。亚细胞定位预测结果发现,Vv4CL01和10蛋白定位于叶绿体上,Vv4CL02、11和14蛋白定位在过氧化物酶体上,Vv4CL03、04和15蛋白定位在质膜上,Vv4CL05蛋白定位于内质网膜上,其余5个成员定位于细胞核中。
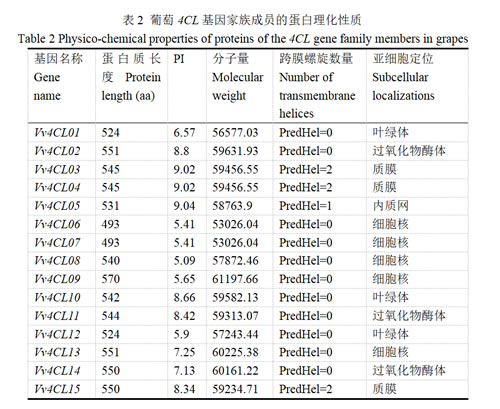
3.2 葡萄4CL家族成员的染色体定位
通过生物信息学方法对葡萄Vv4CL基因家族成员的染色体分布进行系统分析,结果如图1所示,15个成员不均匀分布于7条染色体上。
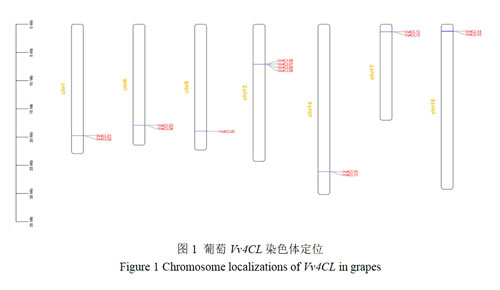
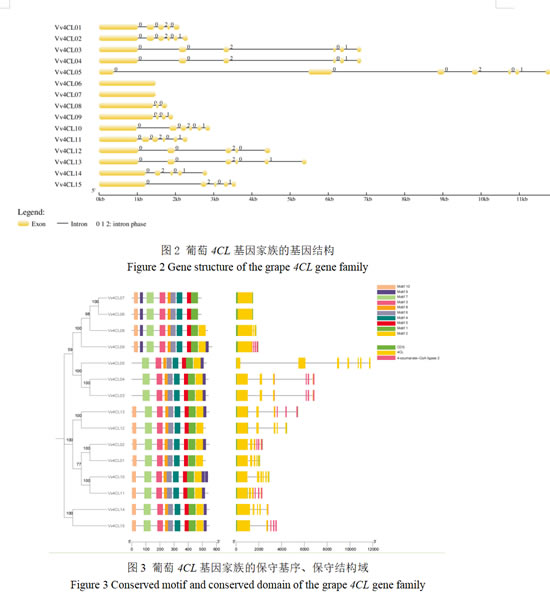
3.3 葡萄4CL家族的基因序列结构、保守基序及结构域分析
如图2所示,葡萄Vv4CL基因家族成员的基因长度范围为1478~11804 bp,其中Vv4CL05最长,Vv4CL06、07最短。不同成员的外显子程度和数目存在显著差异,Vv4CL05的外显子数最多,有7个,但其第1个外显子长度(380 bp)比其他成员(992~1412 bp)显著减少;Vv4CL06和07的外显子数最少,仅有1个;其他大部分成员的外显子数为5~6个。内含子长度和位置呈现高度异质性,Vv4CL05的第1和2个内含子长度分别为5105和2768 bp,Vv4CL03和04的第3个内含子长度均为2728 bp,其他成员的内含子长度相对较短,大多在100-1000 bp。
利用MEME工具鉴定出10个保守Motif(图3),这些功能区域在不同基因中的分布位置和数量有所不同,但总体上保持了一定的保守性。所有成员都含Motif1、3、4、5、6、7和8,体现该基因家族的功能保守性。除Vv4CL03、04和Vv4CL05之外,大部分成员含有Motif10。所有基因CDS长度在1495-1706 bp,均含保守的4CL和4-coumarate-CoA ligase 2功能区域,且两者均位于基因中间部分。
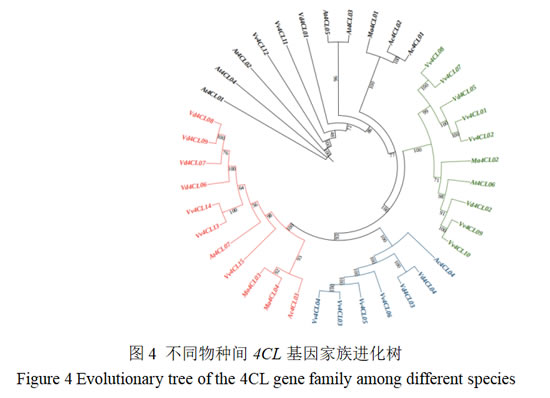
3.4 葡萄4CL基因家族的进化树分析
利用MEGA软件构建葡萄与拟南芥、凤梨、香蕉等物种4CL基因家族系统发育树(图4)显示,不同物种的4CL基因划分为4个主要分支,其中Vv4CL01、02和Vv4CL07~10基因属于第I亚族,和蓝莓Vd4CL02和Vd4CL05、拟南芥的At4CL06及香蕉Ma4CL02归为一类。Vv4CL03~06基因属于第II亚族,和蓝莓Vd4CL03和Vd4CL04聚类紧密。第III亚族含有Vv4CL13~15,同蓝莓Vd4CL06~09、拟南芥At4CL07及香蕉Ma4CL03和Ma4CL04归为一类。Vv4CL11、12基因属于第IV亚族,与其他葡萄4CL基因亲缘关系远,同拟南芥的拟南芥At4CL01~05,归为一类。
3.5 基因顺式作用元件分析
图5显示Vv4CL基因家族启动子激素响应、逆境响应、位点结合和分生组织表达相关的顺式作用元件分布。激素响应相关的顺式作用元件主要6类,包括茉莉酸响应元件(TGACG-motif/CGTCA-motif)、水杨酸响应元件(TCA-element)、脱落酸响应元件(ABRE)、生长素响应元件(AuxRE、TGA-element)和赤霉素响应元件(P-box)。其中,除Vv4CL08~10外,CGTCA-motif/TGACG-motif作用元件广泛分布于多个基因中,Vv4CL13和14最多(7个)。除Vv4CL07和08外,其他基因启动子均含有ABRE元件,Vv4CL12含有6个,Vv4CL01和02仅含有1个。含有TCA-element元件的基因有8个,与水杨酸(SA)响应相关,推测其在葡萄抗病防御机制中发挥作用。AuxRE元件特异性存在于Vv4CL13和14的启动子中。P-box元件仅在Vv4CL12的启动子中检测到,与赤霉素(GA)响应相关,可能参与种子萌发、茎伸长等生长过程。逆境响应相关的顺式作用元件主要有防御和应激响应(TC-rich repeats)、厌氧诱导响应(ARE)、激发子响应(AT-rich sequence)和低温相应(LTR)等4类元件。TC-rich repeats在Vv4CL03-06等基因启动子中均有3个,表明相关基因或参与葡萄抗病原菌感染。除Vv4CL07外,其他基因启动子均含有ARE元件。Vv4CL07、Vv4CL08、Vv4CL11基因启动子中含有AT-rich sequence元件。LTR在Vv4CL09~11、13和14基因启动子中出现,推测与葡萄抗寒性调控相关。此外,仅在Vv4CL07基因启动子中含有1个参与类黄酮生物合成基因调控的MYB结合位点(MBSI),在Vv4CL08基因启动子中发现干旱诱导和光相关的MYB结合位点(分别为MBS和MRE),在Vv4CL01、02和11基因启动子中含有MYBHv1结合位点CCAAT-box元件。
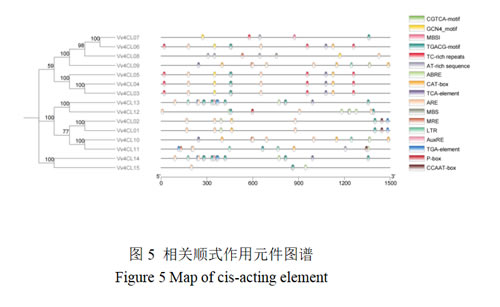
3.6 共线性分析
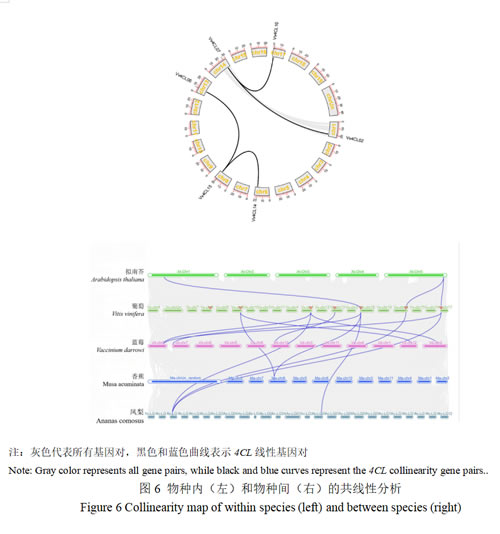
图6所示为葡萄Vv4CL基因家族的共线性可视化图谱。结果显示,Vv4CL基因家族成员在葡萄染色体上呈现片段复制、串联重复的分布特征,共鉴定出3个主要共线性块(Collinear Block)和4对同源基因对,基因对Vv4CL02/Vv4CL07、Vv4CL07/Vv4CL10、Vv4CL06/Vv4CL15、Vv4CL14/Vv4CL15之间存在共线性关系,并被划分到同一亚类(Vv4CL06/Vv4CL15除外),由此可知4CL基因家族在葡萄进化过程中经历了多次基因组复制事件。葡萄与拟南芥、蓝莓、香蕉和凤梨之间的基因共线性分析发现,它们之间的同源基因分别有3对、6对、2对、4对。共线性分析指出葡萄与蓝莓之间的基因对匹配数量远超过葡萄与香蕉之间,这与它们的进化关系相吻合。
3.7 葡萄Vv4CL基因家族在不同组织中的表达模式
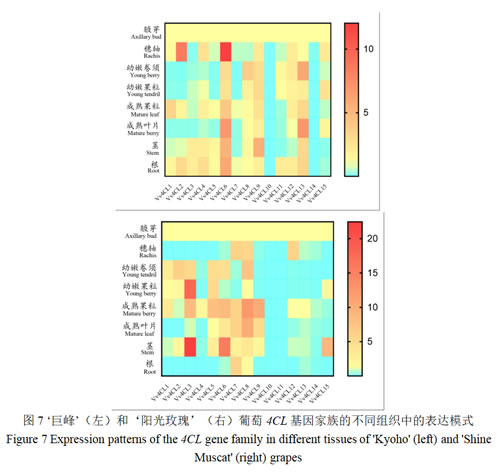
qRT-PCR结果所示(图7),在‘巨峰’葡萄穗轴组织中,Vv4CL2、6的表达量显著高于其他基因;幼嫩卷须中,Vv4CL13的表达量最高;幼嫩果粒中,Vv4CL9、15的表达量较高;成熟果粒中,Vv4CL1、6、13的表达量较高,其他基因表达较一致;成熟叶片中,Vv4CL6、13的表达相对较高;茎中,Vv4CL6、9的表达量较高;根组织中,Vv4CL6的表达量最高,而Vv4CL10、14的表达量最低。Vv4CL10和Vv4CL14在‘巨峰’葡萄中几乎不表达。
在‘阳光玫瑰’葡萄穗轴组织中,Vv4CL7、8、12的表达量较显著,其余几乎不表达;幼嫩卷须中,Vv4CL1~6和Vv4CL8的表达量较一致,其余均较弱;幼嫩果粒中,Vv4CL3的表达量最高,其次是Vv4CL5;成熟果粒中,Vv4CL10~15的表达量不明显;成熟叶片中,Vv4CL8的表达相对较高;茎组织中,Vv4CL3的表达水平最高,其次是Vv4CL6;根组织中,只有在Vv4CL7有显著表达量;Vv4CL10和Vv4CL11在‘阳光玫瑰’葡萄中几乎不表达。
对比分析发现,Vv4CL基因的表达模式在两品种中存在差异。在穗轴中,‘巨峰’中Vv4CL2、6高于‘阳光玫瑰’;而‘阳光玫瑰’的Vv4CL3在幼嫩果粒和茎中的表达量高于‘巨峰’。
4 讨论与结论
4.1 讨论
多个物种的4CL基因家族已被分析研究。本研究在葡萄基因组中成功鉴定出15个4CL家族成员,其数目明显少于汤山酥梨[19](29个),但与毛白杨[20](18个)和石榴[21](12个)大致相同。基因家族在染色体上往往呈不均匀分布以及成簇分布模式[18],染色体定位分析显示Vv4CL基因呈不均匀分布特征,Chr13和Chr18分别富集4个和2个成员,物种内共线性分析发现4对4CL同源基因,表示这种成簇分布模式可能源于串联复制事件。这种复制事件频发,使得葡萄保留了大量同源基因对,其数量远超拟南芥,为葡萄4CL基因家族规模的壮大奠定了基础,同时暗示其在次生代谢途径中具有更复杂的功能分化[22]。
蛋白质保守结构域与系统发育分析可以明确基因家族的进化保守性[23]。本研究中所有4CL成员均携带4CL特征性AMP-binding结构域,其核心基序(Motif1-5)在系统发育相近的成员间高度保守,明确了该家族在催化羟基肉桂酸活化中的基本功能。值得注意的是,Vv4CL02-04、09、11、13和15特有的4-coumarate--CoA ligase 2结构域,可能赋予其底物特异性选择能力,这与拟南芥中At4CL3调控类黄酮合成的功能异曲同工[24]。
基因启动子顺式元件分析中,存在许多抗逆、防御及应激相关的顺式作用元件,与植物抵抗非生物胁迫具有直接或间接的关系[25]。本研究从葡萄4CLs的启动子序列中鉴定到多种与生长发育、激素、逆境等相关的顺式作用元件。CGTCA-motif和TGACG-motif这两种茉莉酸甲酯(MeJA)响应元件广泛分布于多个基因中,表明Vv4CL基因家族的大部分成员可能通过茉莉酸信号通路调控次生代谢产物的合成[21]。AuxRE元件特异性存在于Vv4CL13和14的启动子中,表明该基因可能受生长素调控,参与细胞伸长、分化等生长发育过程[26]。
结构基因的表达常受外界环境对其启动子中顺式作用元件的调控,存在时空差异[25]。在穗轴中,‘巨峰’Vv4CL2和Vv4CL6的高表达与‘阳光玫瑰’中不同,可能导致两个品种穗轴在木质化程度、养分运输效率等方面存在差异,进而影响穗轴的形态和功能,如穗轴的粗细、强度以及对果实的支撑能力等。‘阳光玫瑰’Vv4CL3在幼嫩果粒和茎中的高表达,而‘巨峰’中该基因在这两个部位表达量较低,这可能是造成两个品种果实发育和茎的生长特性差异的重要原因之一。在‘巨峰’的成熟果粒中,Vv4CL3、Vv4CL5~9基因的表达量较高,而在‘阳光玫瑰’葡萄中较低,可能影响果实的大小、形状、口感等品质性状。这些基因表达的特异性可能与两个品种的生长特性、次生代谢物合成及环境适应性密切相关。在生长特性上,可能影响植株的整体形态、生长速度等;在次生代谢物合成方面,可能导致果实和其他组织中次生代谢物种类和含量的差异,进而影响果实的品质和植株的抗逆性;在环境适应性方面,不同的基因表达模式可能使两个品种对不同的土壤、气候等环境条件具有不同的适应能力。
4.2 结论
本研究通过全基因组分析系统鉴定了葡萄中15个4CL基因家族成员,在染色体上呈不均匀分布,系统发育分析将其分为4个亚族,顺式作用元件分析揭示了4CL基因在光响应(G-Box、GT1-motif)、激素调控(茉莉酸、脱落酸、赤霉素响应元件)及逆境响应(低温、氧化胁迫元件)中的多维调控网络。共线性分析进一步表明,葡萄4CL基因家族经历了多次片段复制事件,与蓝莓等物种存在进化保守性,同时具有物种特异性扩张特征。通过qRT-PCR分析‘巨峰’和‘阳光玫瑰’葡萄不同组织中Vv4CL基因家族的表达模式,表明其具有显著组织特异性及品种间差异。本研究结果为进一步探究4CL基因家族参与非生物胁迫的响应提供参考依据,为阐明苯丙烷代谢调控机制提供了重要理论依据。