采后贮运保鲜
李晓雪 鄢骕 袁一林 李桐 张娜 白继新 陈存坤 纪海鹏
摘要:m6A修饰在果蔬成熟软化及采后保鲜中具有重要作用,其生物学功能主要依赖 YTH 结构域蛋白,但目前阳光玫瑰葡萄中的VvYTH尚未见报道。本研究通过生物信息学手段,共鉴定到了阳光玫瑰葡萄中的10个VvYTHs,发现其多数为稳定性较差的亲水酸性蛋白。同时,通过保守基序、保守结构域、基因结构、染色体定位和家族共线性分析发现VvYTHs家族基因的保守性较高。此外,通过转录组及qRT-PCR结果分析发现,VvYTHs参与阳光玫瑰葡萄采后软化进程,其中VvYTH1可能是ClO2处理延缓采后阳光玫瑰葡萄软化进程的关键基因之一。本研究结果不仅可为阳光玫瑰葡萄VvYTHs基因功能的研究奠定坚实的基础,同时也为果蔬保鲜剂的研发提供新的思路。
关键词:阳光玫瑰葡萄;VvYTH基因家族;生物信息学分析;ClO2保鲜处理;基因表达分析
前言
阳光玫瑰葡萄(Vitis labrusca×V. vinifera, Shine Muscat)属葡萄科(Vitaceae Juss.)葡萄属(Vitis L.),由日本杂交育成,于2011年引入我国。因其果实汁多味美,深受消费者喜爱,此外其富含维生素C、维生素K、钾、铁、白藜芦醇和花青素等营养成分,具有较强的抗氧化特性,在抗癌、增强免疫功能、预防慢性疾病的发生等多个方面展现出了巨大的应用潜力。据不完全统计,截止2021年,我国阳光玫瑰葡萄的种植面积和收获面积分别为20806 hm2和13613hm2[1]。尽管我国阳光玫瑰葡萄产量和销售量均位居世界前列,但因采后软化问题造成的葡萄品质下降及腐烂变质是其采后损失的主要因素之一。
二氧化氯(Chlorine dioxide, ClO2)是目前应用较为广泛的食品保鲜剂,因其具有较强的杀菌性及无残留的特性已在2004年被正式批准为果蔬杀菌剂。此外,因其对于维持采后果蔬的品质具有良好的效果而备受关注。2022年,研究发现用ClO2处理阳光玫瑰葡萄后,在贮藏第30天,其硬度下降速率相较于对照组和二氧化硫处理组最小(为35.75 %)。2024年,集贤等]研究发现,低浓度的ClO2缓释剂能够较好的维持红地球葡萄(Vitis vinifera, Red Globe)的硬度。基于此,ClO2对于延缓采后葡萄的软化具有较好的效果,然而对于其背后分子生物学机制的研究,尤其是RNA修饰目前尚处于起步阶段。
RNA修饰是一类在转录后水平上的RNA表观修饰,目前已鉴定出包括mRNA、rRNA、IncRNA和tRNA等在内的各类RNA上的150多种修饰,其中N6-甲基腺嘌呤(N6-methyladenosine, m6A)是真核生物中最常见也是mRNA中丰度最高的可逆甲基化修饰形式。m6A是RNA分子中腺嘌呤核苷(Adenosine, A)N6位置氢原子被甲基取代后形成的一种较为保守的化学修饰。相较于哺乳动物,植物m6A修饰研究起步较晚,植物中的m6A多集中在3' UTR、起始密码子和终止密码子附近,调控mRNA的剪接、稳定性、出核和翻译等,参与植物的生长发育、胁迫响应以及果实的成熟软化等过程。
植物m6A 的生物学功能主要依赖 YTH 结构域蛋白(阅读器,Readers),其是RNA 代谢的主要参与者,在空间、时间和功能上决定着 RNA 的命运。此外,YTHs是目前发现最早且研究较为清楚的 m6A“Reader”,其含有一段由 140个氨基酸组成的区域在其同源蛋白中高度保守,被命名为 YTH(YT521-B homologs)结构域,用于识别 m6A 修饰的碱基。然而,阳光玫瑰葡萄中的VvYTHs基因家族目前尚未有报道。
1 材料与方法
1.1 试验材料
阳光玫瑰葡萄采收自天津市武清区,运回天津市农业科学院国家保鲜工程技术研究中心(天津)冷库。经由0±1 ℃预冷24 h后,挑选出外观完好无机械损伤且成熟度一致的阳光玫瑰葡萄,将其以10±1 kg用微孔膜进行随机分组包装。
分别设置对照组(CK-0:未添加ClO2的初始样品;CK-72d: 未添加ClO2贮藏72天的样品),以及处理组(T3-72d: 添加ClO2的量为2 g/kg贮藏72天的样品;T5-72d: 添加ClO2的量为8 g/kg贮藏72天的样品),每组设置三个生物学重复。
1.2 方法
1.2.1 阳光玫瑰葡萄转录组测序
将CK-0、CK-72d、T3-72d和T5-72d的样品,液氮速冻后,一部分送上海美吉生物有限公司进行转录组测序,另一部分放置于-80 ℃冰箱用于后续qRT-PCR实验。
1.2.2 阳光玫瑰葡萄VvYTH家族成员鉴定
YTH 保守结构域 HMM 文件(PF04146)从 Pfam(https://pfam.xfam.org/)获得,利用 HMMER 3.0 在葡萄转录组数据中检索,得到10个VvYTHs家族成员。
1.2.3 阳光玫瑰葡萄VvYTH家族成员蛋白理化性质分析
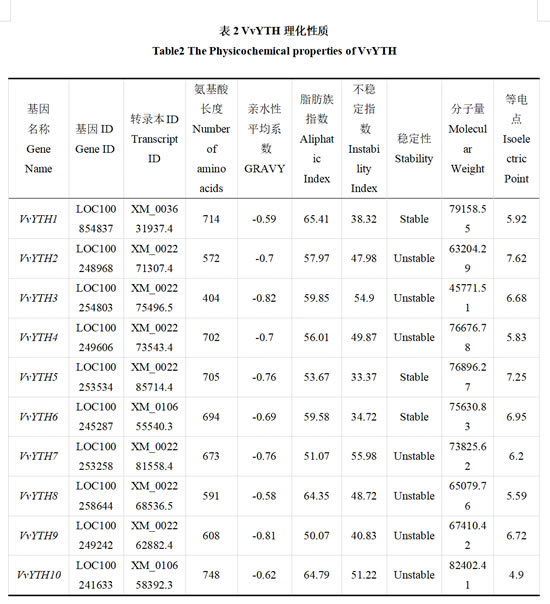
通过在线网站 ExPASy ProtParam (https://web.expasy.org/protparam/)预测10个VvYTHs蛋白的氨基酸序列长度(Amino Acid Length)、分子量(Molecular Weight)、等电点(Isoelectric Point)、亲水性平均系数(GRAVY)、脂肪族指数(Aliphatic Index)及不稳定指数(Instability Index),并基于不稳定指数值判断蛋白稳定性(Instability Index<40 为稳定,Instability Index≥40 为不稳定)。
1.2.4 阳光玫瑰葡萄系统进化树构建和VvYTH家族保守基序、保守结构域分析
在NCBI(https://www.ncbi.nlm.nih.gov/)上下载文献报道过的9个草莓FvYTH蛋白序列,13个拟南芥AtYTH蛋白序列和9个番茄SlYTH蛋白序列[16],与之前筛选到的10个阳光玫瑰葡萄VvYTH1-10蛋白序列采用 MAFFT 7.0(https://mafft.cbrc.jp/alignment/server/index.html)进行多序列比对。之后通过 MEGA 12 用邻接法(neighbor-joining,NJ)构建系统发育树(Bootstrap=1000),并利用在线网站iTOL(https://itol.embl.de/)进行可视化。利用在线网站MEME(http://meme-suite.org/memei)分析10个VvYTH1-10蛋白的保守基序(E-value<1e-5,基序长度 6-50 bp))并可视化。使用在线工具Batch CD-Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)检索其保守结构域,获得hitdata文件,通过TBtools中“Gene Structure View”模块进行可视化展示。
1.2.5 阳光玫瑰葡萄VvYTH家族基因结构及染色体定位分析
物种参考基因组文件及GTF注释文件从NCBI(https://www.ncbi.nlm.nih.gov/)上取得,使用TBtools中“Gene Structure View”模块分析阳光葡萄VvYTH基因家族成员的编码区/非编码区(CDS/UTR)结构。通过TBtools中“Gene Location Visualize from GTF/GFF”模块绘制染色体定位图,标注VvYTH基因在染色体上的物理位置。
1.2.6 阳光玫瑰葡萄VvYTH家族GO富集和蛋白互作分析
通过美吉生物云平台(https://cloud.majorbio.com/)对 VvYTH 蛋白进行 GO 功能注释,采用超几何检验筛选显著富集的条目(P<0.05),并生成气泡图。利用String数据库(https://cn.string-db.org/)对阳光玫瑰葡萄VvYTH家族蛋白序列进行分析,预测其蛋白互作网络,并进行可视化。
1.2.7 阳光玫瑰葡萄VvYTH家族共线性分析和基于转录组数据的表达模式分析
使用TBtools中“One Step MCScanX”模块进行阳光葡萄物种内共线性分析,通过TBtools中“Advanced Circos”工具进行可视化,标注其中的VvYTH家族基因。使用美吉生物云平台(https://cloud.majorbio.com/)基于转录组数据对阳光玫瑰葡萄VvYTH家族在四种不同条件下(CK-0、CK-72d、T3-72d和T5-72d)的表达量进行表达量聚类分析并生成热图。
1.2.8 qRT-PCR 分析VvYTH基因家族成员的基因表达
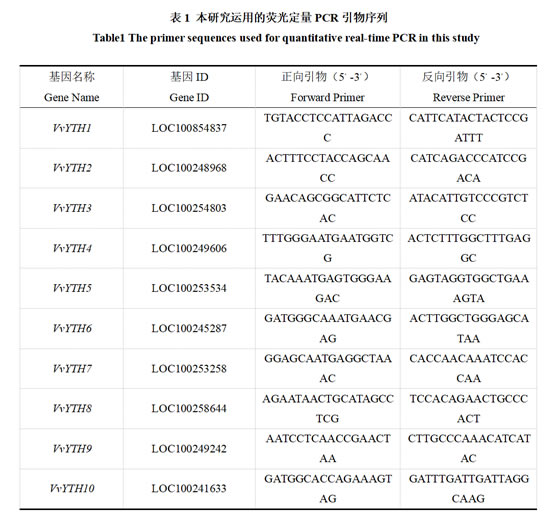
利用Primer Premier 5.0软件对VvYTH基因家族设计引物(表1),内参基因选择VvActin-101(Gene ID:LOC100246825)。实时荧光定量PCR仪采用QuantStudioTM 1 Plus PCR仪,分析软件则是用荧光定量PCR仪自带的QuantStudioTM Design&Analysis SC software v 1.0.0。反应体系按照 TaKaRa Premix Ex Taq TM Ⅱ 的说明书略有改动。反应体系设定为20 μL:cDNA2.0 μL,正向引物和反向引物各 1 μL,TBGreen(TaKaRa Premix Ex Taq TM Ⅱ)10 μL,ddH2O 6 μL。采用两步法PCR反应程序:预变性阶段95 ℃ 30 s;PCR反应阶段95 ℃ 5 s和60 ℃ 30 s,40个循环。试验设置3组平行实验。以VvActin-101为内参基因,用2-ΔΔCt计算VvYTH1-10基因相对表达量。
2 结果与分析
2.1 阳光玫瑰葡萄VvYTH家族成员蛋白理化性质分析
通过HMMER 3.0使用隐马尔可夫模型(Hidden Markov Model,HMM)在转录组数据中检索YTH保守结构域,筛选出了10个阳光玫瑰葡萄VvYTHs家族成员,分别命名为VvYTH1~VvYTH10,对其进行理化性质分析(表2)。其中序列长度最短为404个氨基酸残基,最长为748个氨基酸残基,平均氨基酸残基个数为641。10个VvYTHs的平均等电点为6.366,其中VvYTH2和VvYTH5等电点大于7,呈碱性,而其余VvYTHs等电点均小于7,呈酸性。VvYTH1~VvYTH10的亲水性平均系数(Grand average of hydropathicity,GRAVY)均小于0,标明其均为亲水蛋白。同时,以40为不稳定指数(Instability Index)的界限,可以观察到除VvYTH1、VvYTH5与VvYTH6外,其余VvYTHs蛋白稳定性均较差。
2.2 阳光玫瑰葡萄VvYTH家族系统进化树构建和保守基序、保守结构域分析
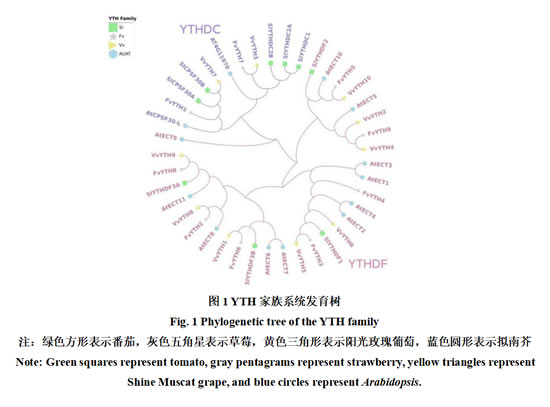
为探究VvYTHs家族与其他物种YTH家族基因的进化关系,对阳光玫瑰葡萄、拟南芥、草莓和番茄共4个物种41个YTHs基因的氨基酸序列进行多重比对并构建系统发育树(图1)。41个蛋白中有11个属于YTHDC亚族,30个属于YTHDF亚族,分别在图中以紫色和粉色表示,其中VvYTH3与FvYTH7亲缘关系较近,VvYTH7与SlCPSF30B亲缘关系较近,表明这两个VvYTH家族成员属于YTHDC亚族,其余VvYTH家族成员属于YTHDF亚族。
为探究VvYTHs家族成员蛋白的保守性,使用MEME和Batch CD-Search分别对其保守基序和保守结构域进行分析,结果显示在阳光玫瑰葡萄VvYTHs家族共鉴定到10个保守基序(Motif1~Motif10)(图2)。同时,该结果符合系统发育分析,亲缘关系相近的家族成员拥有相近的保守基序。YTHDC亚族与YTHDF亚族之间差异较大:属于YTHDF亚族的8个VvYTH家族成员均含有除Motif5、Motif9和Motif10之外的七个保守基序,属于YTHDC亚族的VvYTH3与VvYTH7则仅含有Motif9与Motif10两个保守基序。

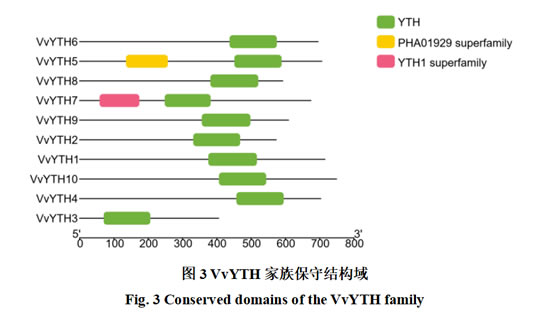
对VvYTHs家族成员蛋白序列的保守结构域的鉴定显示(图3),所有成员均含有YTH保守结构域。除此之外,VvYTH5还存在一个PHA01929超家族结构域,VvYTH7存在一个YTH1超家族结构域。由保守基序和保守结构域分析可知,VvYTHs基因家族属于高度保守的家族。
2.3 阳光玫瑰葡萄VvYTH家族基因结构及染色体定位分析
为进一步探明阳光玫瑰葡萄VvYTHs家族基因的结构特征,利用GTF注释文件绘制VvYTHs基因结构图(图4-A)。结果表明,阳光玫瑰葡萄VvYTHs基因家族均有内含子,外显子数目在6~9个之间。此外,VvYTHs家族基因染色体定位图显示其在染色体上分布均匀(图4-B),10个家族成员随机分布在10条染色体上,每条染色体上均有一个VvYTH基因。其广泛性分布暗示其在多倍体化后的适应性保留,对于阳光玫瑰葡萄复杂性状及其在应对逆境的进化中具有关键性的作用。
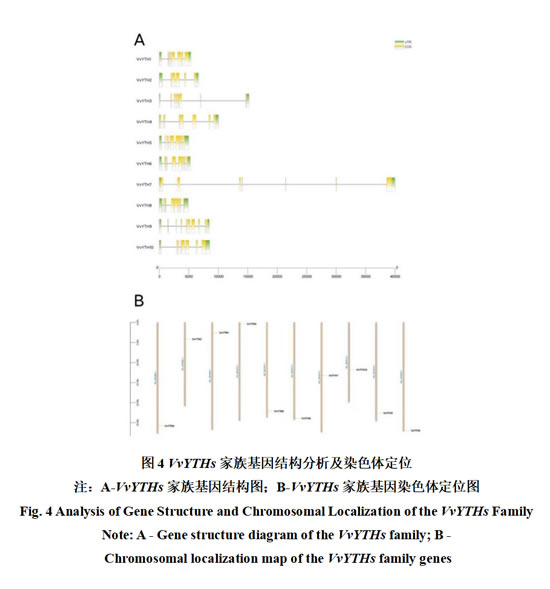
2.4 阳光玫瑰葡萄VvYTH家族共线性分析
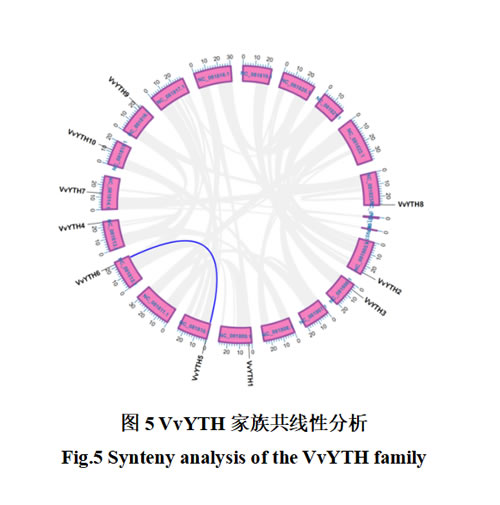
基因复制是新基因产生的基础,推动了基因功能的多样化和生物进化的复杂性,研究表明,基因复制主要通过三种机制促进基因家族的形成和扩展:片段复制(segmental duplication)、串联重复(tandem duplication)和易位事件(transposition events)。其中,片段复制和串联重复被认为是基因家族扩展的主要驱动力。为解析阳光玫瑰葡萄VvYTHs家族基因复制的情况,采用TBtools中“One Step MCScanX”模块进行阳光玫瑰葡萄物种内共线性分析,结果表明VvYTHs基因家族仅有VvYTH5与VvYTH6一对基因存在共线性关系,证明仅存在少量片段复制,同染色体上无密集基因簇,证明无明显串联重复(图5)。
2.5 阳光玫瑰葡萄VvYTH家族GO富集和蛋白互作分析
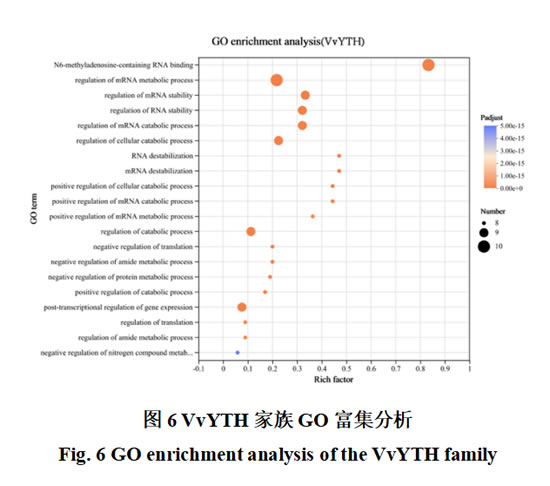
为探究VvYTHs家族蛋白在阳光玫瑰葡萄中发挥的功能与相互作用,对其进行GO功能富集与蛋白互作网络分析。结果表明,VvYTHs 家族主要参与含N6-甲基腺嘌呤的RNA结合(N6-methyladenosine-containing RNA binding)、调节mRNA代谢过程(regulation of mRNA metabolic process)、mRNA稳定性的调节RNA稳定性的调节(regulation of mRNA stability regulation of RNA stability)和mRNA代谢过程的调节细胞代谢过程的调节(regulation of mRNA catabolic processregulation of cellular catabolic process)等生物学过程(图6),表明 VvYTHs 家族在 RNA 加工和降解中具有关键作用,符合前期研究对植物中YTH家族功能的认知。
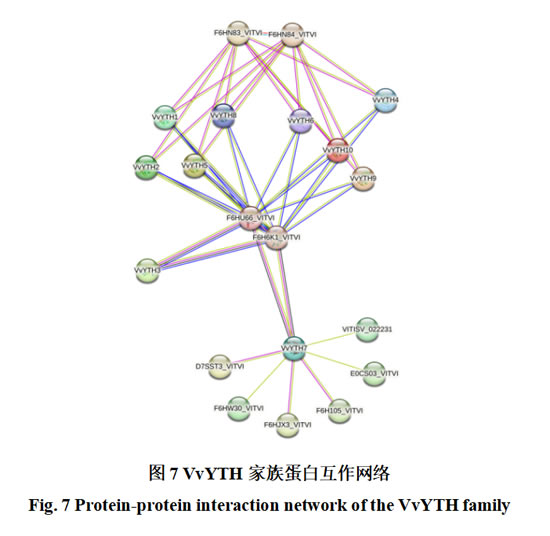
蛋白互作网络的预测是研究基因功能的有效方法之一,为进一步探究VvYTHs的功能,采用String在线工具对其互作网络进行分析。结果显示(图7),VvYTH1~VvYTH10可与多个蛋白互作,其中与VvYTH1~VvYTH10均可互作的F6HU66_VITVI和F6H6K1_VITVI与已知的N6-腺苷甲基转移酶(MT-A70)家族蛋白高度相似,推测其可能参与RNA 的N6-甲基腺苷(m6A)修饰过程,与前人研究相符。
2.6 VvYTH基因表达量分析
在前期研究中,我们发现ClO2处理可延缓采后阳光玫瑰葡萄的软化进程,其中T3-72d处理显著延缓了阳光玫瑰采后软化进程,而T5-72d处理则无此显著效果,为探究阳光玫瑰葡萄VvYTH基因家族在此过程中的作用,对其转录组数据进行了分析(图8)。对CK-0、CK-72d、T3-72d和T5-72d转录组数据进行聚类分析,共得到4个子聚类,这表明VvYTHs基因家族具有多种不同表达模式。VvYTH1、3、5、6和10在贮藏第72天时,表达量相较于CK-0和T3-72上调。VvYTH2、4、7、8和9在贮藏第72天时,表达量相较于CK-0下调。此外,VvYTH1、4、5、6、7、9与10均体现出在T3-72d处理下表达被抑制而在T5-72d处理下表达量回升的现象。表明VvYTH1、5、6和10可能参与了ClO2处理延缓采后阳光玫瑰葡萄软化进程。

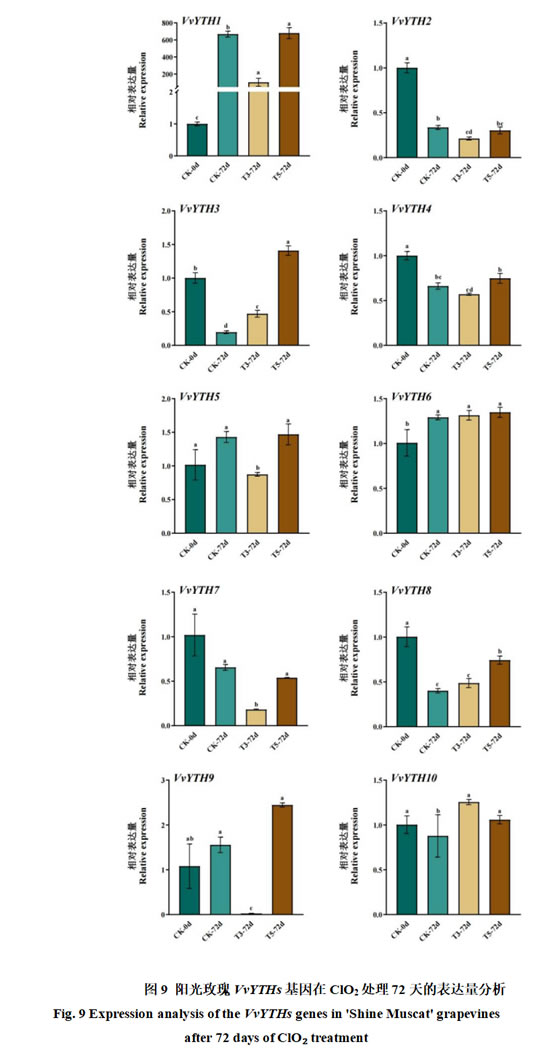
为了进一步验证阳光玫瑰葡萄VvYTHs家族基因在ClO2处理72天后的表达情况,对其进行了qRT-PCR验证(图9)。结果表明,VvYTH2、3、4、6、7、8和10的基因相对表达量较低,VvYTH5和VvYTH9的相对表达量略高,而VvYTH1这个基因成员相比其他基因成员的相对表达量最高,T5-72d的表达量为CK-0组的680倍。VvYTH2、4、7和8的相对表达量在CK-0最高,在CK-72d、T3-72d和T5-72d的条件下,基因相对表达量均有所下降,而VvYTH1和VvYTH6这两个基因与上述基因表达趋势恰好相反,这与转录组中的结果相一致。此外,在T5-72d的条件下VvYTH3、8、9以及10的基因表达水平比CK-72d组显著提高,分别为CK-72d组的7.18、1.85、1.57和1.2倍。同时,VvYTH1和VvYTH5的表达模式较为相近,均为在T3-72d的条件下表达量相较于CK-72d和T5-72d较低,其中VvYTH1的表达量最高。根据转录组与qRT-PCR的结果表明,VvYTH1可能是参与ClO2处理延缓采后阳光玫瑰葡萄软化进程的关键基因之一。
3 讨论与结论
m6A修饰在调控果实成熟、品质维持、逆境响应方面均具有重要作用。2024年,Su等发现m6A修饰参与了呼吸跃变型果实猕猴桃(Actinidia chinensis)的成熟及品质形成,“消码器”AcALKBH10和“阅读器”AcECT9 显著影响了猕猴桃果实可溶性糖和有机酸的积累,对于其果实品质的形成至关重要。Zhou等在国际期刊Nature Plants上发表研究结果,显示番茄m6A去甲基化酶SlALKBH2与硫氧还蛋白还原酶SlNTRC发生互作,SlNTRC能够调控SlALKBH2的氧化还原状态,继而影响其蛋白的稳定性,同时说明SlALKBH2氧化修饰参与了H2O2诱导的果实成熟。然而,截止目前, m6A修饰在延长采后果实保鲜期、抑制采后果实软化进程等方面的研究尚处于起步阶段,且尚未有阳光玫瑰葡萄m6A相关酶(VvYTH)的研究报道。
与动物中的 YTHs 相比,植物YTHs 家族成员较多,在模式植物番茄(Solanum lycopersicum L.)中共鉴定到 9 个,拟南芥(Arabidopsis thaliana L.)中13 个 YTHs(命名为ECT1-12 和 AtCPSF30-L),苹果(Malus pumila Mill.)中 15 个,陆地棉(Gossypium hirsutum L.)中26个],小麦(Triticum aestivum L.)中 39 个。本研究使用隐马尔可夫模型在前期测得的转录组数据中检索YTH保守结构域,共鉴定到了10个阳光玫瑰葡萄VvYTH家族成员,其中VvYTH3和7属于YTHDC亚族,其余属于YTHDF亚族(图1)。通过对VvYTHs理化性质的分析发现,它们均为亲水蛋白,且大部分为蛋白稳定性较差的酸性蛋白(表2),这与陆地棉GhYTHs的蛋白理化性质相类似。
为进一步验证VvYTH家族进化关系的正确性,对其保守基序和保守结构域进行了分析,发现VvYTHs基因家族属于基序和结构域高度保守的家族(图2和3),与番茄中的SlYTHs基因家族类似,推测其功能上也相对保守。在染色体定位层面,番茄的9个SlYTHs基因分布在5条染色体上,黄瓜(Cucumis sativus L.)的5个CsYTHs分布在3条染色体上,而阳光玫瑰葡萄的10个VvYTHs基因随机分布在10条染色体上(图4-B),表明其在多倍体化后的适应性保留,对于其功能的多样性具有一定的作用。同时,通过分析阳光玫瑰葡萄物种内共线性,发现VvYTHs中仅有VvYTH5与VvYTH6一对基因存在共线性关系(图5),该结果与陆地棉GhYTHs的共线性关系不同,表明VvYTHs基因家族的进化可能未涉及染色体片段复制事件。
植物YTHs 主要通过识别 m6A 修饰调控 RNA 代谢,继而参与植物的生长发育、果实成熟衰老、逆境响应等。为探明VvYTHs基因的功能,结合课题组前期测得的转录组数据(即CK-0、CK-72d、T3-72d和T5-72d)对其进行GO富集分析和蛋白互作分析(图6和图7)。结果表明,VvYTHs主要参与RNA代谢并且与N6-腺苷甲基转移酶(MT-A70)家族蛋白高度相似的F6HU66_VITVI和F6H6K1_VITVI互作,这与番茄、猕猴桃、燕麦中YTH的主要功能一致。ClO2处理是果蔬采后保鲜的常用手段,其可以有效杀灭病原微生物同时延缓采后葡萄的软化进程。Bian等研究发现,番茄m6A“阅读器”SlYTH2通过液-液相分离形成蛋白-RNA凝聚体,抑制靶标mRNA的翻译,继而影响番茄的香气,同时在敲除SlYTH2后,番茄的货架期显著延长。本研究通过分析转录组数据(即CK-0、CK-72d、T3-72d和T5-72d)中VvYTHs的表达量(图8),发现VvYTH1、5、6和10可能参与了ClO2处理延缓采后阳光玫瑰葡萄软化进程。同时,利用qRT-PCR对转录组数据进行了验证(图9),结果表明低浓度ClO2处理可显著抑制VvYTH1在贮藏72天时的表达量,表明其可能是ClO2处理延缓采后阳光玫瑰葡萄软化进程的关键基因之一。
综上所述,本研究利用生物信息学手段,共筛选到了10个阳光玫瑰葡萄VvYTHs基因,对其进行了理化性质、进化关系、保守基序、保守结构域、基因结构、染色体定位、家族共线性分析、GO富集、蛋白互作、转录组及qRT-PCR分析,初步探明了VvYTHs在进化及功能层面具有一定的保守性,VvYTH1可能是ClO2处理延缓采后阳光玫瑰葡萄软化进程的关键基因之一。该研究结果可为阳光玫瑰葡萄RNA表观修饰研究提供研究思路,同时也为以m6A修饰为靶点的果蔬保鲜剂的研发奠定基础。