酿酒葡萄品种改良岗位
摘要:本研究以酿酒葡萄‘赤霞珠’和‘西拉’为亲本,对其正反交519株杂交后代展开研究,旨在揭示果实不同部位酚类物质遗传变异规律,为酿酒葡萄杂交育种提供理论支撑。研究结果显示正反交后代果实酚类物质在果皮中均低于亲中值,表现为衰退变异,果肉和种子中均值都高于亲中值,遗传倾向于酚类指标含量高的亲本,总体表现为超亲现象明显,表现出一定的杂种优势。研究明确了‘赤霞珠’与‘西拉’正反交后代果实不同部位酚类的遗传倾向,为酿酒葡萄杂交亲本的科学选配和新品种选育提供了重要参考。
酿酒葡萄人工杂交是有目的的控制花粉和选择亲本的品种来获得杂交种子,国内酿酒葡萄育种进展我国早在20世纪50年代初就开始葡萄杂交育种工作,自 21世纪以来,育种工作进入了快速发展时期,葡萄品种的选育数量逐年增加。
葡萄果实酚类物质也是酿造葡萄酒的重要影响因子。它不仅赋予葡萄酒各种颜色而且使葡萄酒具有各种复杂的口感和味感特征。研究发现,在优良的酿酒品种的选择与适宜的酿造工艺的确定过程中,酚类物质特性具有重要的作用。黄酮类化合物是种类繁多的化合物,无论它们的存在与否,它们都对葡萄酒的品质有很大的贡献。虽然葡萄酒中的类黄酮含量和成分在一定程度上反映了葡萄酒酿造的过程,但主要的决定因素是采收时葡萄的品质。因此,已经进行了大量研究,以了解葡萄中黄酮类化合物的性质,影响其生物合成的因素,以及如何将这些知识用于收获时管理和操纵浆果中黄酮类化合物的成分。在葡萄中,酚类化合物(主要是类黄酮)是葡萄酒显色的条件之一,花青素(负责红酒的色泽特征的彩色颜料),单宁(负责红酒颜色的长期稳定性)和黄酮醇(可能通过色素沉着影响酒色的化合物)也起到很大的作用。此外,类黄酮也会影响其他感官特性,例如涩味,苦味和酒体等。在果实成熟期时,并非是完全成熟时酚类化合物达到峰值,而是在采收前几天。全面了解葡萄的成分对于了解酿酒过程和生产更高品质的葡萄酒至关重要。葡萄果实中含有很多酚类活性物质,尤其是黄酮化合物,主要存在于葡萄果皮和葡萄籽中,包括总黄酮、原花色素、黄烷醇、花色苷等,在酒精发酵过程进入酒体,对红葡萄酒的色泽、口感、香气、抗氧化、抗菌有重要作用。酚类物质含量主要受葡萄品种、栽培环境和酿造工艺的影响,其中最重要的决定因素是葡萄品种。
本研究以‘赤霞珠’与‘西拉’正反交后代为试验材料,进行果实成熟期只要酚类物质遗传倾向分析,阐明其遗传变异规律,一方面可以为酿酒葡萄杂交育种亲本选择提供重要的参考依据,另一方面选育出的优良后代可作为中间或终端优系应用于生产,显著改良品种应用结构。
1 材料与方法
1.1 试验材料
研究材料来源于山西农业大学果树研究所酿酒葡萄育种圃,地理位置为北纬37 度 23分,东经 112 度 32 分。年平均气温 10.6℃,降水量为 400~600 mm,无霜期 160~180 d,海拔 800 m,年日照时数 2300 h,有效积温3675℃,土壤为砂壤及粉砂壤土,土壤 pH 值 7.8。
研究所用试材为‘赤霞珠’ב西拉’(正交)、‘西拉’ב赤霞珠’(反交)两个正反交组合的 F1代。通过真假子代的筛选以正交215个后代和反交 304 个后代的真子代为试验材料。
1.2 试验方法
1.2.1 指标测定的方法
对从超低温取出的葡萄果实进行果皮、果肉、种子的分离,用滤纸将果皮和种子上的汁液吸收干净后取2 g果皮、6 g果肉、2 g种子分别于100 mL棕色容量瓶中,用70%的乙醇于暗处浸提24 h,过滤后的滤液为酚类物质提取液。采用Folin-Ciocalteu法测定总酚和单宁含量;通过氯化铝比色法测定总类黄酮;采用正丁醇-盐酸比色法测定原花色素含量;称0.4 g果皮于100 mL棕色容量瓶中,用1%盐酸-甲醇溶液于暗处提取12 h后滤液为总花色苷提取液,采用 pH 值示差法测定花色苷含量,以二甲花翠素葡萄糖苷表示( mg·g-1 FW)。
1.2.2 数据统计分析方法
通过 Excel 对数据进行整理,利用相关计算公式计算变异系数、组合传递力、优势率、广义遗传力等遗传指标,用 Excel 对统计的每个性状的遗传规律。
变异系数、组合传递力、优势率等的计算公式如下。
变异系数: CV/%=S/F×100;
组合传递力(遗传传递力): Ta/% = F/P × 100;
优势率:Ha/% = (F–P)/P × 100;
利用方差分析计算广义遗传力:
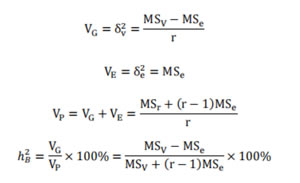
S:标准差;F:后代平均值;P亲本平均值(亲中值);r 随机区组设计中的重复次数;VG:基因型变量;VP:表型变量;VE:环境变量;ℎ�2:广义遗传力。
2 结果与分析
2.1‘赤霞珠’和‘西拉’正反交后代果实不同部位总酚含量的遗传
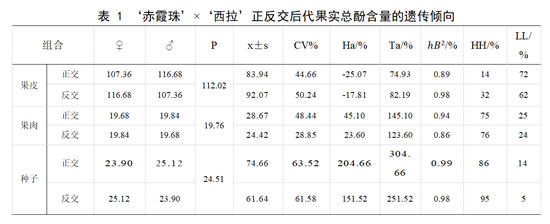
亲本‘赤霞珠’果皮总酚含量平均值为 107.36 mg·g-1,果肉总酚含量均值为19.68 mg·g-1,种子总酚含量均值为23.90 mg·g-1;亲本‘西拉’果皮总酚含量为116.68 mg·g-1,果肉总酚含量 19.84 mg·g-1,种子总酚含量为 25.12 mg·g-1。
在正交后代果皮中总酚含量的平均值低于亲中值,超低亲率72%,遗传有趋于低含量母本‘赤霞珠’的趋势,优势率为负值,杂种优势不明显。组合传递力为74.93%低于100%,变异系数为 44.66%,存在较广的分离。正交后代果肉总酚含量平均值高于亲中值,介于双亲19.68 ~19.84 mg·g-1 范围内的概率为0,超高亲率占75%,优势率为正值 45.10%,具有一定的杂种优势。组合传递力高于100%,母本变异传递给后代的能力较强。变异系数 48.44%,存在较广的分离。正交后代种子总酚含量的平均值高于亲中值,优势率高于 100%。组合传递力高于100%。总酚的超高亲率最高达 86%,变异系数为 63.52%。反交后代果皮中总酚含量的平均值低于亲中值,优势率为负值。组合传递力低于100%。超低亲率占62%,变异系数为50.24%。果皮中总酚含量的偏度小于1。在果肉和种子中,总酚含量的平均值高于亲中值,优势率分别为23.60%和151.52%,种子的总酚含量具有明显的强亲效应,组合传递力分为123.60%和251.52%。超高亲率均高于超低亲率,具有一定的杂种优势。反交后代的果肉和种子中,总酚含量的广义遗传力为 0.864 和 0.987,变异受环境影响小,主要来自遗传效应。
3.2 ‘赤霞珠’和‘西拉’正反交后代果实不同部位单宁含量的遗传
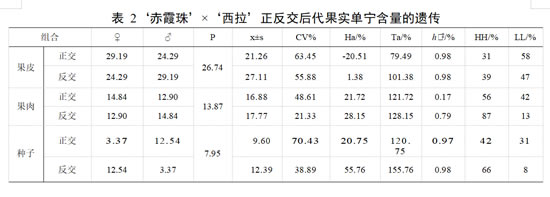
亲本‘赤霞珠’果皮单宁含量平均值为29.19 mg·g-1,果肉单宁含量平均值为 14.84 mg·g-1,种子单宁含量平均值为 3.37 mg·g-1;亲本‘西拉’果皮单宁含量平均值为 24.29 mg·g-1,果肉单宁含量平均值为 12.90 mg·g-1,种子单宁含量平均值为 12.54 mg·g-1。正交后代果皮单宁含量的平均值低于亲中值,优势率为负值,表现为衰退变异。组合传递力中低于 100%,超低亲率 58%,单宁的变异系数为 63.45%。正交后代果肉和种子中单宁含量的平均值均高于亲中值,优势率均为正值。具有一定的杂种优势。组合传递力均高于 100%,正交组合遗传能力强。果肉和种子中单宁含量介于双亲间的植株极少,超高亲率均高于超低亲率,表现为超高亲遗传。单宁含量在果肉和种子中的变异系数高,存在较广的分离。
正交后代中果肉中单宁含量遗传的广义遗传力为 0.174,变异受环境影响较大。 反交后代果皮中单宁含量的平均值与亲中值相近,优势率 1.38%,组合传递力高于101.38%。超低亲率占 47%,变异系数为 55.88%。反交后代果肉和种子中,单宁含量的平均值高于亲中值,优势率为 28.15%和 55.76%,有一定的杂种优势。组合传递力 128.15%和 155.76%。果肉中单宁含量介于双亲之间的概率为 0,超高亲率均较高,表现明显的杂种优势。果肉中单宁含量的偏度小于 0,整体呈左偏分布或左偏态,个体分布向小于平均数的方向偏斜,有利于对单宁较少品种的选择。
3.3 ‘赤霞珠’和‘西拉’正反交后代果实不同部位总黄酮含量的遗传
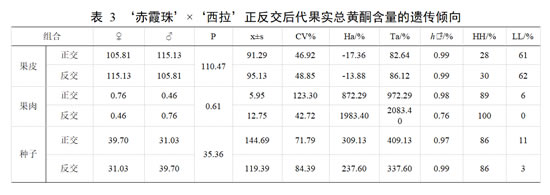
亲本‘赤霞珠’果皮总黄酮含量平均值为 105.81 mg·g -1,果肉总黄酮含量平均值为0.76 mg·g-1,种子总黄酮含量平均值为 39.70 mg·g-1;亲本‘西拉’果皮总黄酮含量平均值为 115.13 mg·g -1,果肉总黄酮含量平均值为 0.46 mg·g -1,种子总黄酮含量平均值为31.03 mg·g -1。正交后代果皮中总黄酮含量的平均值低于亲中值,优势率为负值,不具有杂种优势。组合传递力为 82.64%,超低亲率高于超高亲率,变异系数为 46.92%,遗传趋于总黄酮含量低的亲本遗传。在正交后代果肉和种子中,总黄酮含量的平均值均高于亲中值,优势率均较高,表现为强亲效应。组合传递力高于 100%,超高亲率均在 80%以上,遗传具有明显的母本优势。反交后代果皮总黄酮含量的平均值低于亲中值。超高亲率占 30%低于超低亲率 62%,但有一定的杂种优势。总黄酮含量组合传递力 86.12%,变异系数为 48.85%,存在较广的分离。反交后代果肉和种子中,总黄酮含量的平均值高于亲中值,存在一定的加性效应,超高亲率均高于超低亲率,且果肉超高亲率 100%,杂种优势明显。
3.4 ‘赤霞珠’和‘西拉’正反交后代果实不同部位黄烷醇含量的遗传
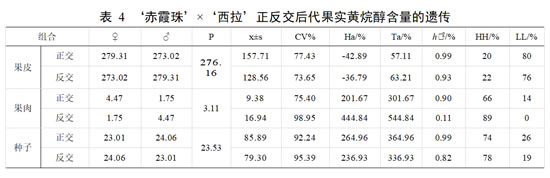
亲本‘赤霞珠’果皮黄烷醇含量均值为 279.31 mg·g-1,果肉黄烷醇含量均值为 4.47 mg·g -1,种子黄烷醇含量均值为 23.01 mg·g -1;亲本‘西拉’果皮黄烷醇含量均值为273.02 mg·g-1,果肉黄烷醇含量均值为 1.75 mg·g-1,种子黄烷醇含量均值为 24.06 mg·g-1。
正交后代果皮黄烷醇含量的平均值低于亲中值,优势率为负值,不具有杂种优势。黄烷醇含量在果皮中相比在果肉和种子中组合传递力较低,超低亲率 80%,表现为衰退趋势。变异系数为77.43%,存在较广的分离。在正交后代果肉和种子中,黄烷醇的平均值高于亲中值,存在一定的加性效应,遗传倾向于黄烷醇含量高的亲本。优势率高,表现为强亲效应。在果肉和种子中,黄烷醇含量超高亲率均较高,表现为明显的杂种优势。两者变异系数均较高,存在较广的分离。
反交后代果皮中黄烷醇含量的平均值低于亲中值,超低亲率76%,优势率为负值,遗传倾向于黄烷醇含量低的亲本,呈现衰退变异。组合传递力 63.21%,变异系数为73.65% ,呈广泛分离。反交后代果肉和种子中黄烷醇含量的平均值均高于亲中值,优势率均高于100%,表现为强亲效应。组合传递力均较高,表明反交组合在果肉和种子中黄烷醇含量的遗传能力强。超高亲率占 89%,黄烷醇变异系数均较大,分离广泛。其中果肉中黄烷醇含量遗传的广义遗传力为 0.11,变异受环境影响较大。
3.5 ‘赤霞珠’和‘西拉’正反交后代果实不同部位原花色素含量的遗传
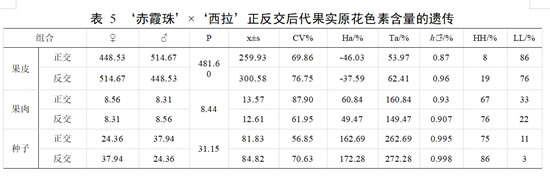
亲本‘赤霞珠 ’果皮原花色素含量平均值为 448.53 mg·g-1,果肉原花色素含量平均值为 8.56 mg·g-1 ,种子原花色素含量平均值为 24.36 mg·g-1;亲本‘西拉 ’果皮原花色 素含量平均值为 514.67 mg·g-1,果肉原花色素含量平均值为 8.31 mg·g-1 ,种子原花色素含量平均值为 37.94 mg·g-1。正交后代果皮酚类物质含量中原花色素的平均值低于亲中值,优势率为负值。组合传递力低于 100%,超低亲率高于超高亲率,变异系数为 69.86%。在正交后代果肉和种子中,原花色素含量的平均值均高于亲中值,优势率为正值,具有明显的杂种优势。组合传递力均高于 100%,正交组合的遗传能力强。两者在原花色素含量中均存在超亲遗传,且超高亲率明显,遗传趋于果肉和种子中原花色素含量高的亲本。
反交后代果皮中原花色素含量的平均值低于亲中值,优势率为负值,超低亲率占 76%,遗传倾向于原花色素含量低的亲本,表现为衰退变异。变异系数为 76.75%,分离较广泛。反交后代的果肉和种子中,原花色素含量的平均值均高于亲中值,优势率为正值,超高亲率均高于超低亲率,遗传倾向于原花色素含量高的亲本,有一定的加性效应。果皮、果肉和种子原花色素含量的广义遗传力均在 0.90 以上,变异主要来自遗传效应,受环境影响小。
3 结论
本研究围绕‘赤霞珠’与‘西拉’正反交后代果实不同部位酚类物质,分析其遗传规律,为酿酒葡萄育种提供了关键参考。在正反交后代果皮的5种酚类指标中,仅反交后代单宁含量具有一定的杂种优势,其余4个指标总酚、总黄酮、黄烷醇和原花色素含量在正交和反交遗传中均表现为衰退变异。在后代果肉酚类物质遗传中,单宁、总黄酮、黄烷醇和原花色素含量主要倾向于‘赤霞珠’;在后代种子酚类物质遗传中,总酚、单宁、黄烷醇和原花色素含量主要倾向于‘西拉’。在正反交后代遗传中均出现了超亲遗传现象,为育种者新品种的选育提供一定的参考价值。