苗木生产岗位
卢浩成 于可济 王军
摘 要:本研究旨在阐明行向引起的微气候变化啊如何改变赤霞珠葡萄和葡萄酒的香气物质。选择了四个行向的葡萄园,并测定来自不同叶幕侧的葡萄和葡萄酒。结果显示,与曝光的侧面相比,遮光减少了果穗周围的总辐射和高温胁迫的时间。葡萄初级代谢物受果穗位置的影响较小。草本植物的香气在遮光侧的葡萄和葡萄酒中浓度更高。温度较高的叶幕一侧加速了葡萄中萜类化合物的挥发但并增加了降异戊二烯的水平。东北-西南葡萄园在口感和香气感官评分方面表现出最显著的差异。在相关性网络中,香蕉、菠萝和草莓的气味与最终的感官评分高度相关。本研究为葡萄园行向选择、分区收获和葡萄酒的定向生产提供了潜在的策略。
关键词:微气候;代谢组;转录组;香气;感官
葡萄园行向是葡萄园内自然微气候变化的最明显因素之一,因为它直接导致叶幕两侧不均一的阳光吸收和环境温度(Hunter et al., 2020)。一些研究强调了葡萄园行向在不同树冠侧修改葡萄和酒的成分中的关键作用(Gao et al., 2021; Hunter & Volschenk, 2018; Wang et al., 2022)。一旦葡萄园确定了某种行向,便无法轻易改变,树冠两侧之间固有的微气候异质性也会持续存在。然而,可以采取一些解决方案来减少树冠两侧之间的微气候异质性,例如在遮光侧增加基部摘叶量(Gambacorta et al., 2022)。此外,从不同叶幕两侧进行分开(分区)收获也是一种潜在策略,尽管关于这种方法的报告较少。
挥发性香气物质是葡萄中的重要的次级代谢物,可以被视为酒的风味指纹,代表特定品种、特定酒区甚至特定葡萄园的特征。随着分析技术的发展,越来越多的挥发物在葡萄和葡萄酒中被识别,为“挥发物组学”的研究提供了充足的化学基础(Lu et al., 2023b)。然而,挥发物的数量庞大且其形成机制复杂,使得理解它们如何对生物和非生物胁迫作出反应成为一项挑战。挥发性化合物通常可以作为抗氧化剂,保护植物免受非生物胁迫下产生的活性氧物质的有害影响(Wang et al., 2020),而由于叶幕两侧经历不同的胁迫水平,挥发性香气物质会受到葡萄园行向的影响。例如,Wang等(2022)观察到,在东西向行的葡萄园中,葡萄中的香气物质更容易受到影响,与相应的微气候异质性较大相关,而与南北向葡萄园相比则表现不同。与这些结果不同的是,Gao等(2021)发现,每个行向中两侧叶幕果穗的香气物质差异不大,并且与年份相关。尽管现有研究已报告了行向对不同树冠侧浆果成分的显著影响,但这些差异是否也适用于从分开收获(每侧分开)获得的葡萄酒仍然是一个值得探究的问题。迄今为止,关于赤霞珠葡萄及其葡萄酒的挥发物组学和相关感官特征的研究仍然有限,特别是在不同行向和果穗位置的情况下。更重要的是,将挥发物组学发现与最终的感官感知联系起来,也是风味相关研究的关键目标。
全球变暖现象确实对葡萄酒行业带来了变化,许多传统地区的葡萄园面临比以往更多的热胁迫(Gutiérrez-Gamboa et al., 2021)。在中国,超过70%的葡萄种植区和葡萄酒产量集中在西北炎热地区,如新疆、宁夏和甘肃(Lu et al., 2023b)。在这些地区,夏季频繁出现热浪,因此葡萄种植者采用浓密树冠策略,以保护葡萄免受晒伤和高温的不利影响(Lu et al., 2022)。然而,这种做法导致特定行内各个树冠之间的隔离更加明显。即使在可能具有更均衡光分布的南北向葡萄园中,面朝西侧的树冠在下午也遭受比东侧更多的热胁迫(Lu et al., 2021)。因此,该地区的葡萄园为评估不同行向下树冠侧之间葡萄挥发物组学的差异提供了良好的机会。我们假设,由行向造成的树冠侧之间的微气候变异可能会影响侧分开葡萄酒的典型香气特征。为验证这一假设,我们选择了新疆地区四个不同行向的商业葡萄园,评估了来自两个树冠侧的葡萄挥发性代谢物成分以及这些葡萄酿成的酒的感官特征。我们的目标是提供关于葡萄和葡萄酒挥发物组学与感官感知及聚集位置之间关系的知识。我们的结果将有助于在炎热地区制定有根据的葡萄园建设策略,特别是涉及分区采收的策略。
1 材料与方法
1.1 田间试验设计
田间实验在新疆玛纳斯地区(北纬44°24′,东经86°26′,平均海拔522 m)的四个商业葡萄园进行。玛纳斯产区属于半干旱的炎热产区,该地区葡萄生长季内的活动积温可达2200 °C(Wang et al., 2021; Jones et al., 2010)。这四个葡萄园分别定义为南北向(NS,北偏东25°)、东北-西南向(NE-SW,北偏东49°)、西北-东南向(NW-SE,北偏西45°)和东西向(EW,北偏东85°)。所有葡萄园均于2009年至2011年间建立,种植品种为赤霞珠。各个葡萄园采取了一致的管理措施,行距固定为1 m × 3 m,采用改良后的VSP整形方式。每株葡萄修剪为17-23个芽,保留27-33个果穗。所有葡萄园均统一实施产量控制策略,产量控制为9-12 t/ha。葡萄在每两周进行一次滴灌,从5月持续到8月,每次灌溉量为750 m³·ha⁻¹。在每个葡萄园内,选择了三行相邻的均匀葡萄进行实验。每个样本组由三组50株连续的葡萄组成,均位于其中一行。使用HOBO微型气象站(Onset,Bourne,MA,USA)进行果穗区微气候的调查,以精确监测总辐射和温度。数据收集持续三个生长季,从开花期(5月底)开始,到收获期(9月中旬)结束,测定为每五分钟一次。
1.2 样品的采集
在2019-2021年三个连续的生长季节中,分别在四个行向的不同叶幕侧对浆果进行采样,采样覆盖葡萄发育的整个阶段。采用E-L系统作为葡萄物候期的判定并作为采样依据(Coombe, 1995)。在以下阶段采集浆果:2019和2021年:E-L 33、E-L 35、E-L 36、E-L 37、E-L 38;2020年:E-L 33、E-L 35、E-L 37、E-L 38。每次采样每个重复随机采集400粒浆果,来自果穗的不同部位,包括顶部、中部和底部。采集的浆果立即用液氮冷冻,并在-80 °C的冰箱中保存以备后续分析。
1.3 小规模发酵
当葡萄的可溶性固形物含量达到23-25 °Brix时,进行小规模发酵,每个处理采收60 kg果穗(共3个生物学重复,每个生物学重复20 kg)。人工破碎除梗后入罐,同时加入20 mL 4% H2SO3和0.4 g果胶酶。入罐24 h后添加4 g活化好的Lalvin D254酵母(Lu et al., 2023a)。干净纱布盖住发酵罐口,每天早晚压帽两次,控制酒精发酵温度在22 °C左右,并测定发酵醪比重和温度。酒精发酵结束后(比重降至0.998以下且维持2 d不再变化)分离皮渣,自流汁和压榨汁移至10 L密闭玻璃容器,添加乳酸菌进行苹-乳发酵,水封隔绝氧气,控制发酵温度为20 °C左右。苹-乳发酵结束后加入12 mL 6%的H2SO3并装瓶打塞,置于酒厂酒窖进行瓶储。
1.4 葡萄中的初级代谢产物的测定
每个重复样本随机选取80 g浆果,破碎后置于离心机6000 × g离心5分钟,并通过0.22 μm的聚醚砜过滤器过滤。采用Agilent 1200系列高效液相色谱仪(Agilent Technologies, Inc., Santa Clara, CA, USA)测定糖和有机酸,使用HPX-87H Aminex离子交换柱(300 × 7.8 mm,Bio-Rad Laboratories, Hercules, CA, USA)分离化合物(Zhang et al., 2022a)。流动相为5 mmol/L H2SO4,流速设置为0.6 mL/min。糖使用Agilent G1362A折光指数检测器检测。有机酸使用Agilent G1315D可变波长检测器检测,检测波长为214 nm。所有样本的注入体积为20 μL,柱温为60°C。糖类通过外标物葡萄糖和果糖定性和定量。有机酸则通过酒石酸、苹果酸和柠檬酸为外标物。
1.5 葡萄和葡萄酒中挥发性化合物的测定
葡萄汁和葡萄酒中的游离香气化合物直接测定,结合态的香气物质按照Lu等(2022)的方法提取。利用气质联用的方法分析样品中的香气物质。载气为高纯氦气,流速为1 mL/min。升温程序如下:50°C保持1 min,然后以3°C/min升温至220°C,最后保持5 min。其余条件如下:进样口温度为250°C,葡萄酒的检测采取5:1分流模式,葡萄汁的检测采取不分流模式,电离方式为电子电离(electron ionization, EI)源,电离能为70 ev,离子源温度为230 °C,质谱接口温度为280 °C,质量扫描范围为30~350 u。挥发性香气物质的定性根据NIST标准谱库中的保留指数和质谱信息进行分析。香气物质的定量根据对应的标准曲线进行。
1.6 转录组分析
2019年从NS和EW葡萄园收集的葡萄样品用于转录组分析。每个重复(共3个重复)取20粒葡萄去籽,并在液氮保护下研磨成粉末。总RNA从粉末中提取,采用Spectrum™植物总RNA试剂盒(Sigma-Aldrich, Carlsbad, CA, USA)(Chen等,2017年)。RNA的质量和完整性使用NanoDrop 2000(Thermo Fisher Scientific, Waltham, MA, USA)和Agilent 2100/LabChip(Agilent Technologies, Inc., Santa Clara, CA, USA)进行评估。根据之前的研究(Yin et al., 2022),构建并使用Agilent 2100生物分析仪和Thermo AB StepOnePlus RT-qPCR系统(Thermo Fisher Scientific, Waltham, MA, USA)检查cDNA文库。随后使用Illumina NovaSeq 6000测序平台(Illumina, San Diego, USA)在PE150模式下进行测序。过滤后的序列比对至参考基因组Vitis vinifera - PN40024基因组V4。KEGG富集分析用于筛选与挥发性化合物途径相关的DEGs。本研究的转录组数据已上传至国家基因组科学数据中心(NGDC),登录号为CRA013750 (https://ngdc.cncb.ac.cn/gsa/browse/CRA013750)。
1.7 感官评价
在葡萄酒发酵完成并装瓶两个月后,按照全球葡萄酒中国鉴评体系的规定进行感官品评(Lu et al., 2023)。该评鉴体系包含四个部分:色泽(10分)、香气(30分)、口感(50分)和总体评价(10分)。来自当地酒厂的10名经验丰富的评估员参加了感官分析,包括3名女性和7名男性,年龄在23至48岁之间。所有品评员均自愿参加本实验,并遵循1975年赫尔辛基宣言的原则。本研究的感官评估获得中国农业大学研究伦理委员会批准,参考编号为CAUHR-20220711。训练和品评程序根据Lu等(2023)的方法进行。
1.8 统计分析
数据处理使用Microsoft Excel 2019软件进行,显著性分析通过SPSS 22.0完成,显著性水平为p < 0.05(Duncan多重检验或t检验)。图表生成使用GraphPad Prism 8.0.2和Microsoft Excel 2019软件,包含柱状图、折线图和雷达图。主成分分析和正交偏最小二乘-判别分析使用SIMCA 14.1软件进行和可视化。相关网络和热图使用OmicShare进行绘制(https://www.omicshare.com/tools)。。
2 结果与讨论
2.1 微气候数据与葡萄汁中主要初级代谢物的分析
通过评估果穗区的微气候条件,不同果穗位置的总辐射(SR)表现出明显差异(图1)。此外,NS、NE-SW和NW-SE葡萄园的西侧以及EW葡萄园的南侧经历了更严重的高温胁迫,表现为高温天数的增加。
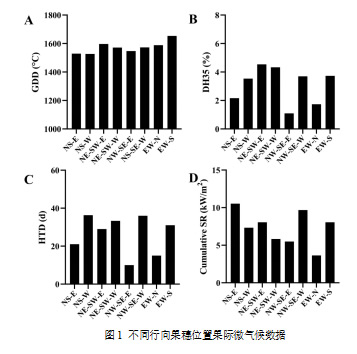
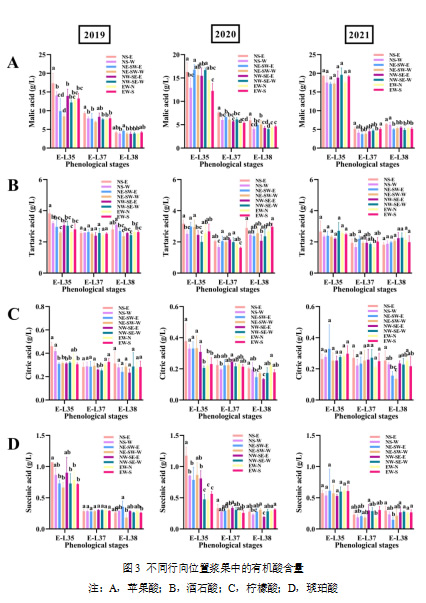
注:A,从开花期到采收期的活动积温(以10 °C为基准计算);B,从开花期到采收期高温小时数占比;C,从开花期到采收期的高温天数(每日最高温度超过35°C);D,从开花期到采收期的累计总辐射在葡萄成熟过程中,不同行向葡萄园中的糖积累基本一致,如图2所示。显著差异(p < 0.05)仅出现在2019年,与其他葡萄园相比,NS葡萄园的果实中果糖和葡萄糖浓度更高,而这一现象在随后的年份未再出现。同一葡萄园的两侧叶幕果实的差异较小。有机酸方面(图3),2019和2020年,NS-E侧的果实中苹果酸和酒石酸浓度显著高于NS-W侧(p < 0.05)。2020年,柠檬酸和琥珀酸在NE-SW和NS葡萄园中也有差异,而其他年份未见此现象。在四个行向葡萄园中,只有NS-E的苹果酸和酒石酸浓度显著高于NS-W侧(p < 0.05)。这可能是由于西侧的强烈热胁迫,较高温度加速了葡萄中有机酸的降解(Sweetman et al., 2014年)。值得注意的是,在所有葡萄园中,果实均从同一葡萄藤的两侧叶幕采集,这些冠层并非隔离的部分。葡萄藤内物质的运输是正常的,尤其是糖分。糖由叶肉细胞合成,并通过韧皮部从叶片运输至果实。因此,即使曝光的一侧吸收了更多阳光且具有较高的光合能力,所产生的蔗糖仍然可以向另一侧冠层的果穗迁移(Keller, 2015)。同样,其他溶质如有机酸、氨基氮化合物和各种次级代谢物也能通过韧皮部移动(Iland et al., 2011)。尽管如此,行向对有机酸的影响似乎比对糖的影响更为显著,这可能是由于大多数果实中的苹果酸和酒石酸直接在果实中合成,因此对微气候变化更为敏感(Sweetman et al., 2009)。
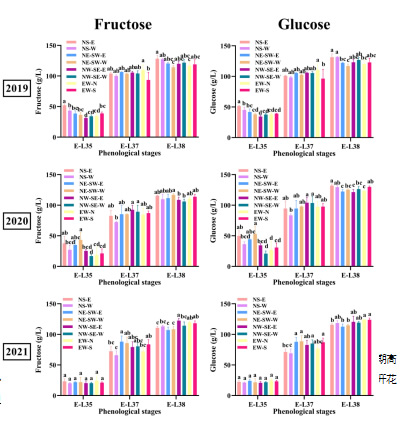
图2 不同行向位置浆果中的葡萄糖和果糖含量
注:A,从开花期到采收期的活动积温(以10 °C为基准计算);B,从开花期到采收期高温小时数占比;C,从开花期到采收期的高温天数(每日最高温度超过35°C);D,从开花期到采收期的累计总辐射
2.2 行向和果穗位置对葡萄香气的影响
2.2.1 C6/C9类化合物
通过t检验筛选出了受行向和果穗位置影响显著的几种C6/C9化合物(图4A):己烯醛、(E)-2-己烯醛、1-己醇、(Z)-3-己烯-1-醇和(E)-2-己烯-1-醇。己烯醛和(E)-2-己烯醛是赤霞珠葡萄中最丰富的醛类物质,在EW葡萄园中表现出显著差异。EW-N葡萄的己醛和(E)-2-己烯醛浓度显著高于EW-S,且至少在两个生长季节中保持这一差异。转录组数据表明,四个VviLOX基因(Vitvi01g01562、Vitvi05g00472、Vitvi14g00235、Vitvi14g00246)为脂氧合酶途径中差异表达的基因(DEGs),其中两个基因(Vitvi01g01562、Vitvi05g00472)在早期阶段(E-L 33,E-L 35)在EW-S葡萄中表达下调。然而,2019年己烯醛和(E)-2-己烯醛并未表现出显著差异(图2B)。在E-L 38阶段,相较于对应的另一侧,VviHPL基因(Vitvi12g00405)在EW-S和NS-W葡萄中上调。脂氧合酶途径下游,C6/C9醛类通过醇脱氢酶的作用转化为C6/C9醇类。在赤霞珠葡萄中,1-己醇是所有C6/C9醇中浓度最高的。NE-SW葡萄园中,2019年和2020年东侧葡萄的1-己醇浓度高于西侧。在NW-SE葡萄园中,2019年NW-SE-E的1-己醇浓度低于NW-SE-W,而2020年则出现了相反的结果。除1-己醇外,EW-N葡萄的(Z)-3-己烯-1-醇浓度也显著高于EW-S。六个VviADH基因被选为NS和EW葡萄园中的DEGs。两个基因(Vitvi04g01670、Vitvi18g01223)在EW-S葡萄的后期发育阶段(E-L 37,E-L 38)相较于EW-N上调,而VviADH基因(Vitvi07g03083)在开花期初(E-L 35)在EW-N葡萄中下调。与EW-S葡萄中两种上调的VviADH基因一致,EW-S葡萄在E-L 37后表现出更高的1-己醇浓度,尽管该差异未达到显著性(p < 0.05)。相反,在采收时,EW-N葡萄的(Z)-3-己烯-1-醇浓度高于EW-S,而在前期发育阶段未出现这一差异。在NS葡萄园中,五个VviADH基因(Vitvi04g01670、Vitvi04g01671、Vitvi07g04173、Vitvi18g01223、Vitvi18g01226)在NS-W葡萄中相较于NS-E都表现出下调。在E-L 35阶段,NS-W葡萄的(Z)-3-己烯-1-醇浓度低于NS-E,但这一趋势在E-L 37阶段发生了逆转,采收时两者未表现出显著差异。
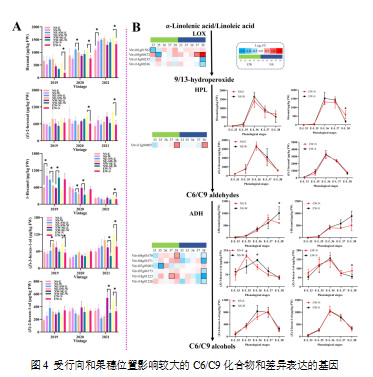
注:(A)采收时差异调控的C6/C9化合物;(B)脂氧合酶途径中的差异表达基因及其在发育阶段中对应的C6/C9化合物的浓度变化。热图以EW-S与EW-N、NS-W与NS-E的对比计算的log2倍数变化值进行可视化表示。具有粗边框的框表示NS-E与NS-W或EW-N与EW-S之间的差异表达基因。图中的“*”表示在某一行向排列的葡萄园内,叶幕两侧葡萄之间的显著差异(p < 0.05,t检验)。
2.2.2 萜类化合物
尽管赤霞珠葡萄被认为是一种中性品种,萜类化合物浓度相对较低,但这些化合物的低阈值仍可能对最终葡萄酒中的愉悦香气有所贡献(Song et al., 2023)。萜类化合物的合成对气候条件敏感,总体萜类化合物浓度在半干旱地区呈一致的下降趋势,这一点得到了类似气候条件下其他研究的支持(Zhang et al., 2016)。与C6/C9化合物类似,EW葡萄园的萜类化合物在采收时表现出更大的变化(图5)。2019年和2021年,EW-N葡萄的β-环柠檬醛浓度显著高于EW-S葡萄(p < 0.05)。相比之下,EW-S葡萄的1-萜品醇(结合态)浓度显著高于EW-N葡萄(p < 0.05)。在NW-SE葡萄园,NW-SE-W的1-萜品醇(结合态)浓度显著高于东侧(p < 0.05),2019年和2020年呈现出相同的结果。通过分析萜类生物合成途径中的DEGs,发现2019年NS葡萄园中的DEGs数量多于EW葡萄园。在EW葡萄园中,VviHMGS(Vitvi14g04271)在EW-S葡萄中上调,而VviHMGR3(Vitvi03g00262)和VviDXS1(Vitvi05g00372)在EW-S葡萄中的表达在E-L 33和E-L 36阶段分别下调。在葡萄发育的后期阶段,EW-N和EW-S葡萄中的d-柠檬烯、β-环柠檬醛、α-卡拉可烯和1-萜品醇(结合态)存在显著差异(p < 0.05),且只有1-萜品醇(结合态)在EW-S葡萄中更高。与主要的合成期(转色前)不同,萜类化合物的变化在降解期(E-L 36至E-L 38)中得到了检测(Zhang et al., 2016)。迄今为止,萜类化合物的降解机制尚不明确。作为抗氧化剂,萜类化合物可以对抗反应性氧种(ROS),并增强葡萄藤对高光照和高温等非生物胁迫的抗性,尽管这样会导致含量的降低(Gutiérrez-Gamboa et al., 2021;Wang et al., 2020)。在NS葡萄园,NS-W葡萄在采收时(E-L 38)表现出比NS-E葡萄更高的游离形式d-柠檬烯和β-环柠檬醛浓度(p < 0.05),但在E-L 37阶段表现出较低的游离形式芳樟醇和顺式芳樟醇氧化物浓度(p < 0.05)。在NS葡萄园的16个DEGs中,5个基因下调,10个基因上调。在NS-W葡萄中,只有VviHMGR1(Vitvi17g00241)在E-L 36阶段下调,并在E-L 38阶段上调,较NS-E葡萄有所不同。参与萜类生物合成的关键基因,如VviDXRs和VviDXSs(Gutiérrez-Gamboa et al., 2021),在NS-W葡萄中上调,这与代谢物的变化趋势一致。从NS和EW葡萄园的结果来看,相对曝光的位置,如NS-W和EW-S,表现出较低的游离形式萜类化合物浓度,而其对应的遮阴位置则相对较高。结合态的萜类化合物则不太受行向的影响,因为只有1-萜品醇(结合态)为差异调控萜类化合物。具有糖苷结构的结合态萜类化合物在水解或酸水解之前不会挥发,从而抑制了它们在非生物胁迫下的释放。
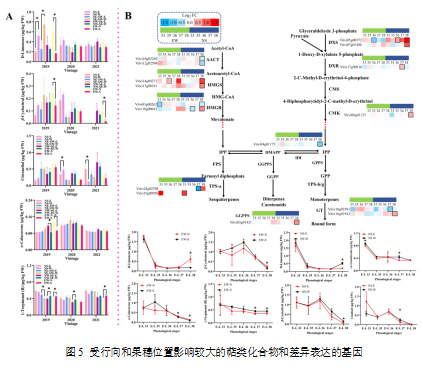
注:(A)采收时差异调控的萜类化合物;(B)萜烯代谢途径中的差异表达基因及其在发育阶段中对应的化合物的浓度变化。热图以EW-S与EW-N、NS-W与NS-E的对比计算的log2倍数变化值进行可视化表示。具有粗边框的框表示NS-E与NS-W或EW-N与EW-S之间的差异表达基因。图中的“*”表示在某一行向排列的葡萄园内,叶幕两侧葡萄之间的显著差异(p < 0.05,t检验)。
2.2.3 降异戊二烯
降异戊二烯来源于类胡萝卜素的降解,这一过程涉及一个复杂的生物合成途径,其基本机制尚未完全理解。至今,负责类胡萝卜素特异性氧化降解和进一步形成降异戊二烯的关键酶尚未被深入研究。类胡萝卜素二氧化酶(CCDs)被确定为参与降异戊二烯合成的关键酶(Schwab et al., 2008)。在采收时,降异戊二烯的变化在NW-SE葡萄园中尤为显著,特别是在2019年和2021年(图6)。NW-SE-W葡萄的vitispirane A、vitispirane A(结合态)、TDN(1,1,6-trimetil-1,2-dihydronaphthalene,结合态)和TPB((E)-1-(2,3,6-trimethylphenyl)buta-1,3-diene,结合态)浓度均显著高于NW-SE-E葡萄(p < 0.05)。这表明,葡萄中的降异戊二烯在曝光且温度较高的果穗位置浓度更高。同样,Cain等(2018)证明,转色后升高的温度促进了葡萄中类胡萝卜素的降解。此外,Scherzinger等(2008)显示,高温胁迫(38 °C)会导致CCDs的表达增加。在NS葡萄园中,采收的葡萄差异较小,只有在2019年,NS-E葡萄中的TPB(结合态)浓度显著高于NS-W葡萄(p < 0.05)。在EW葡萄园,EW-S葡萄在2019年采收时显示出较高(p < 0.05)的vitispirane A和TDN(B)浓度,而在其他年份未观察到这种现象。对于NE-SW葡萄园,2021年出现显著差异(p < 0.05),西侧的葡萄在vitispirane A和vitispirane B(结合态)浓度上均高于东侧葡萄。通过类胡萝卜素生物合成途径,在EW葡萄园中,只有VviZEP2(Vitvi13g01817)发生了差异表达,在EW-S葡萄中下调,相较于EW-N葡萄。然而,在2019年葡萄发育过程中,EW-S葡萄在E-L 33、35阶段的vitispirane A浓度显著高于对应的EW-N葡萄(p < 0.05)。在NS葡萄园中,15个基因发生了差异表达,其中8个基因在NS-E葡萄中上调,如VviPSYs(Vitvi04g02011,Vitvi12g00084)。在代谢物方面,NS-E葡萄显示出较高(p < 0.05)的6-甲基-5-庚烯-2-酮(结合态)浓度,同时其结合态的柠檬烯酮浓度较NS-W葡萄显著较低(p < 0.05)。
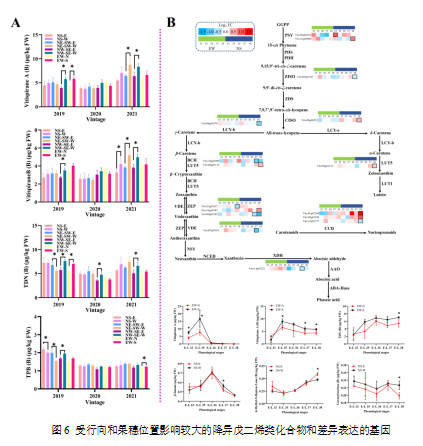
注:(A)采收时差异调控的降异戊二烯类化合物;(B)降异戊二烯代谢途径中的差异表达基因及其在发育阶段中对应的化合物的浓度变化。热图以EW-S与EW-N、NS-W与NS-E的对比计算的log2倍数变化值进行可视化表示。具有粗边框的框表示NS-E与NS-W或EW-N与EW-S之间的差异表达基因。图中的“*”表示在某一行向排列的葡萄园内,叶幕两侧葡萄之间的显著差异(p < 0.05,t检验)。
2.3 行向和果穗位置对葡萄酒香气的影响
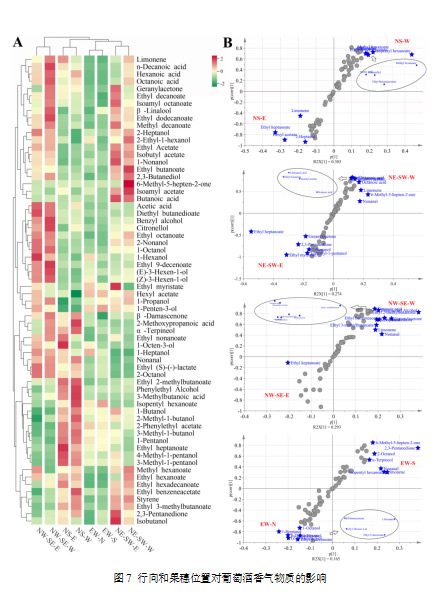
尽管葡萄组成分析可以为酿酒提供指导,但从葡萄到葡萄酒香气的转变是一个复杂的过程,涉及酵母有或没有参与的生物化学代谢,因此必须评估最终葡萄酒中的挥发性化合物。为了筛选受行向和果穗位置影响的关键挥发性物质,采用了聚类分析和正交偏最小二乘法判别分析(图7)。C6醇,如1-己醇、(E)-3-己烯-1-醇和(Z)-3-己烯-1-醇,在NW-SE葡萄酒中浓度最高。NS方向的葡萄园生产的葡萄酒具有最高浓度的高阶醇,如3-甲基-1-丁醇、2-甲基-1-丁醇和1-戊醇,超过了其他葡萄园葡萄酒中的浓度。NE-SW葡萄园的葡萄酒则具有较高的乙酸酯类物质,如乙酸乙酯、异丁酸乙酯和异戊酸乙酯。此外,OPLS-DA被用来选择每个葡萄园中由葡萄架侧面调控的差异香气。在NS葡萄园中,NS-E葡萄酒中的柠檬烯浓度高于NS-W葡萄酒,但香茅醇浓度较低。乙酸庚酯是NS、NE-SW和NW-SE葡萄园中的共同标志性化合物,且在东侧葡萄架上的葡萄酒浓度高于西侧。在NW-SE葡萄园中,大多数通过OPLS-DA选择的标志性化合物在NW-SE-W葡萄酒中的浓度高于NW-SE-E葡萄酒,如香叶烯酮、α-松油醇和柠檬烯。在EW葡萄园中,EW-N葡萄酒的1-己醇、(Z)-3-己烯-1-醇和β-大马士革酮浓度较高,而柠檬烯、α-松油醇和6-甲基-5-庚烯-2-酮的浓度则较低。EW-N葡萄酒中C6醇浓度较高的发现与本实验中葡萄的结果以及之前的研究(Scafidi et al., 2013)一致。因此,从葡萄到葡萄酒的研究结果表明,更阴凉的葡萄藤架有利于热区生产的赤霞珠葡萄酒中绿叶挥发物的积累。然而,之前的报告也发现,紫外线辐射减弱会降低浆果中(E)-2-己烯醛的浓度(Liu et al., 2015)。因此,C6化合物生物合成的调控是复杂的,可能依赖于品种、微气候和年份(Bureau et al., 2000)。
2.4 行向和果穗位置对葡萄酒感官品评的影响
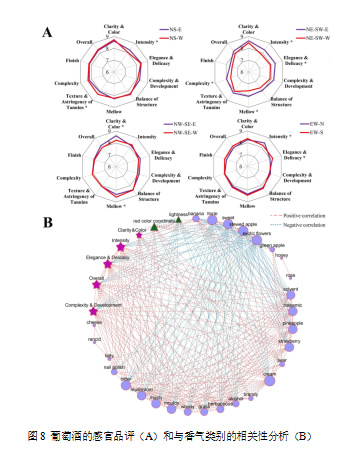
图8展示了来自四种行向葡萄园的葡萄酒的感官结果。在所有葡萄园中,NS、NE-SW和EW葡萄园的香气方面差异显著(p < 0.05)。在NS葡萄园中,NS-E葡萄酒的香气强度得分高于NS-W葡萄酒,但其口感和单宁强度得分较低。NE-SW-E葡萄酒的香气和口感方面得分均高于NE-SW-W葡萄酒,且香气强度、优雅细腻度、口感圆润和复杂度的得分达到了显著水平(p < 0.05),表明针对性分区采收有助于酿酒生产。在NW-SE葡萄园中,香气在感官分析中差异较小,而NW-SE-E葡萄酒在颜色上得分较高(p < 0.05),但口感圆润度得分较低。在EW葡萄园中,EW-N葡萄酒的香气强度得分较高,但优雅细腻度的得分低于EW-S葡萄酒。值得注意的是,受到较少高温胁迫的果穗位置,如NS-E、NE-SW-E和EW-N,香气强度得分较高,表明减少葡萄生长期的高温有助于防止葡萄酒香气的降解(Gutiérrez-Gamboa等,2021)。
将感官结果与挥发性化合物的气味活性值进行相关性网络分析(图8B)。葡萄酒的澄清度和颜色的评分与红色色调呈正相关,而与亮度值呈负相关。在所有的香气类别中,花香、甜香、溶剂香、香脂香、菠萝香、草莓香、奶油香都与其他节点表现出较高的连接性,表明它们在网络中作为核心香气的重要性。香蕉、菠萝、草莓、奶油、香脂和溶剂香气都与香气相关的三个感官方面以及葡萄酒的整体评定呈正相关。然而,由β-大马士酮带来的典型香气,如花香、甜香、炖苹果香和异国花香,都与感官参数呈负相关。如Pineau等(2007)所示,β-大马士酮的阈值范围较广,取决于使用的基质,其对红酒的贡献可能是间接的而非直接的。因此,在本研究中,β-大马士酮所带来的典型香气未能很好地贡献于感官结果似乎是合理的。类似地,Nicolli等(2020)发现,β-大马士酮与赤霞珠葡萄酒的香气强度、红香和干果香的相关性较弱。然而,β-大马士酮与其他挥发性化合物之间可能发生相互作用,不仅仅作为香气增强剂,这改变了其在红酒中的典型气味并导致了检测阈值的不确定性。除了与水果相关的香气外,通常被认为是不愉快的香气,如溶剂香、香脂香和奶油香,也与香气强度、优雅和复杂性呈正相关。因此,在评估它们对葡萄酒的贡献时,考虑其浓度也非常重要。例如,较高的醇类物质是葡萄酒中典型溶剂香的来源,如果浓度控制在300 mg/L以内,它们可以贡献于葡萄酒香气的复杂性(Swiegers & Pretorius, 2005)。
3 结论
不同葡萄园行向和果穗位置造成了微气候的显著差异,但葡萄藤中物质的运输缩小了葡萄中初级代谢产物的差异,糖类的变化比有机酸更为明显。葡萄中品种香的变化与每个行向的光照和热量分布有关。受到高温胁迫的叶幕侧在整个生长季所经历的高温天数更多,这加速了萜类化合物的挥发和降解,同时提高了葡萄中降异戊二烯的浓度。来自东西行向葡萄园北侧的葡萄酒中β-大马士酮浓度比南侧高出13%-32%。与草本植物相关的香气在葡萄和葡萄酒中的变化一致,并且在遮光的一侧叶幕果实中浓度较低。水果相关的香气在感官结果中发挥了核心作用,香蕉、菠萝和草莓的香气与香气感官评分高度相关(R2 > 0.7)。