鲜食葡萄品种改良岗位
1 材料与方法
1.1 材料
供试鲜食葡萄品种为‘玫瑰香’(MGX)、‘瑞都科美’(RDKM)、‘瑞都红玉’(RDHY)和‘瑞都香玉’(RDXY),于2017和2018年9月采收于北京市林业果树科学院葡萄试验园。试验地点在北京市林业果树科学研究院冷库(2 ± 1 ℃,,90% RH)。
1.2 试验方法
选择颗粒饱满、果粒大小均匀、无病、无机械伤、无落粒的葡萄果穗放入开孔纸板箱,每箱5 kg。预冷至-1 ~ 0 ℃,放入PE葡萄保鲜膜中,封口入库。每个品种设3-5次重复,每隔15天(d)进行调查并取样测试相关指标。
1.3 外观品质指标测定
失重率:穗重失重率(%)=(贮前穗重-贮后穗重)/贮前穗重×100%;烂果率:烂果率(%)=(腐烂果粒的个数/果粒总数)× 100%;果柄耐拉力:将葡萄果柄与弹簧秤相连,沿果粒纵轴方向拉至果柄从果实上脱离,记录弹簧秤最终读数,即为果柄耐拉力,每个处理重复10-20次以上,取平均值;可溶性固形物采用手持式折光仪PAL-1(Atago Co,Tokyo,Japan)测定;可滴定酸采用0.1 mol·L-1 NaOH 进行测定,含量以酒石酸表示,3 次重复。
1.4 单萜化合物提取和测定
单萜的提取和测定参考Wen等2015年发表的方法,略有改动。首先葡萄果实样品去除种子,以避免对挥发性化合物的提取产生任何可能的影响。其余果实部分(约50 g)在液氮中研磨,研磨过程中加入1 g PVPP和0.5 g D-葡萄糖酸内酯。得到的果粉样品在4 ℃低温浸渍4 h后,立即在4 ℃下以8000×g离心10 min,得到澄清的葡萄汁。澄清葡萄汁直接检测游离单萜化合物。对每个样品进行三次独立提取。
糖苷结合态化合物提取,固相萃取柱(Cleanert PEP-SPE柱,天津博纳艾杰尔公司)依次经10 mL甲醇和10 mL水活化后,加入2 mL上述澄清葡萄汁样品。经2 mL水洗脱去除一些糖、酸等低分子量的极性化合物后,加入5 mL二氯甲烷洗脱进一步去除大部分游离态香气物质的干扰,最后用20 mL甲醇将结合态香气物质洗脱,收集至50 mL的圆底烧瓶内,整个固相萃取过程洗脱剂流速保持2 mL/min。所得甲醇洗脱液在旋转蒸发器下蒸发至干燥,然后重新溶解在10 mL的2 M柠檬酸-磷酸盐缓冲溶液(pH 5.0)中。在40 ℃的培养箱中,用200 μL 糖苷酶AR 2000(100 mg/L,在pH 5.0的2 M柠檬酸盐/磷酸盐缓冲液中)酶解结合型挥发性化合物16小时。每个样品做两个独立的重复。
此后,在以下SPEME条件下提取游离和结合态形式挥发物:将5 mL提取液与10 μL 4-甲基-2-戊醇(内标)和1g NaCl混合在20 mL聚四氟乙烯硅隔膜盖小瓶中。将样品瓶在40 ℃下在500 rpm搅拌下平衡30分钟。之后将活化的SPME针头(Supelco,Bellefonte,PA,USA)插入小瓶顶空,在40 ℃下吸附挥发分30 min。最后,将SPME针头插入GC进样口8 min,释放挥发物。
气相色谱与质谱联用仪(GC-MS)型号:Agilent 7890B GC和Agilent 5977A MS (Agilent,美国)。毛细管柱为HP-INNOwax 60m×0.25mm×0.25μm (J&W Scientific,美国)。
GC-MS条件参考Wu等[16]发表的方法:载气为高纯氦气(He,>99.999%),流速为1 mL/min;进样口温度为250 ℃,采样不分流模式,解析时间8 min;升温程序为50 ℃保持1 min,然后以3 ℃/min 升温到220 ℃,保持5 min。质谱电离方式为EI,离子源温度为230 ℃,电离能为70 ev,四级杆温度为150 ℃,质谱接口温度为280 ℃,质量扫描范围为30-350 u。
数据处理和分析是通过ChemStation软件(安捷伦科技公司)进行。利用NIST 05标准库中参考标准品的保留指数和质谱匹配来鉴别挥发性化合物。当标准品不可用时,处理保留指数与文献中报告的保留指数的比较。为了量化,使用标准物与内标物的峰面积比与参考标准物浓度对有参考标准物的挥发物进行量化,而使用具有相似碳原子或结构的标准物对没有可用标准物的挥发物进行量化。
1.5 RNA -Seq和加权基因共表达网络分析
使用分别在贮藏后0、15、30天收集的RDHY、RDXY和MGX葡萄进行RNA-Seq分析。总RNA提取和质量分析参考我们之前描述的方法进行(Wang et al., 2022)。使用Illumina his eq 2500基于Illumina标准程序构建了27个来自收集的样品的测序文库(每个时间点3个生物重复)并进行测序,该程序在Origingene公司(https://www.origin-gene.com/)。
生成的原始数据(原始读数)首先通过内部Perl脚本进行处理。通过从原始数据中去除包含接头的读数、包含poly-N的读数和低质量读数获得高质量数据。此外,计算数据的Q20、Q30、GC含量和序列重复水平。然后使用HISAT2(Kim et al., 2019)比对到参考基因组序列(https://plants.ensembl.org/Vitis_vinifera/Info/Index)。基因表达水平的定量通过每百万个作图片段的每千个转录物碱基的片段来估计(FPKM,Trapnellet al., 2010)。利用EdgeR软件进行各组样本之间的DEGs分析(Robinson et al., 2010)。使用R包WGCNA模块(Langfelder & Horvath, 2008)进行自动网络构建功能。
1.6 RT-qPCR验证
为了验证转录组数据,使用SYBR qPCR Master Mix(天根生物有限公司)和CFX96实时PCR检测系统(Bio-Rad)进行了qRT-PCR分析。所有检测基因的引物均由来自NCBI的Primer-Blast(https://www.ncbi.nlm.nih.gov/tools/primerblast/)。两个VvGAPDH(CB975242)和VvUbiquitin(EC 929411)用作内参基因。
1.7 数据处理与统计分析
数据统计分析利用分析软件SPSS 13.0,采用Duncan 多重比较进行显著性方差分析,最低显著水平P<0.05;主成分分析和聚类分析采用MetaboAnalyst 4.0;绘图采用Excel 和Sigma Plot 10.0。
2 结果
2.1 两个葡萄品种低温贮藏过程中果实理化和贮藏特性变化
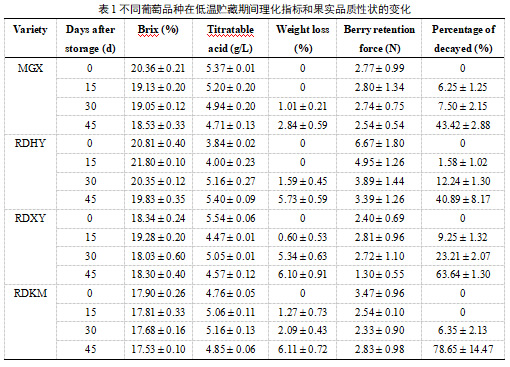
表1显示包括可溶性固形物含量、可滴定酸、失重率、烂果率和果柄耐拉力等基本生理指标。结果表明,贮藏30天后,四个葡萄品种的可溶性固形物含量均呈下降趋势。而4个品种果实中可滴定酸含量略有不同。‘玫瑰香’(MGX)的可滴定酸含量在贮藏至45天时逐渐下降;低温贮藏期间‘RDHY’的可滴定酸含量增加。在贮藏后期(贮藏后30天),四个葡萄品种的果实重量损失迅速增加。与其他两个品种相比,‘MGX’和‘RDHY’的果实显示出较低的重量损失。贮藏期间,4个品种浆果的果柄耐拉力均下降,RDHY的果柄耐拉力明显高于其他3个品种。四个葡萄品种在贮藏后45天都表现出较高的腐烂率;‘MGX’和‘RDHY’的腐烂率低于其他两个品种。综合来看,‘RDHY’和‘MGX’在低温贮藏期间的表现优于‘RDXY’和‘RDKM’。
2.2 四个品种葡萄果实中的单萜化合物组分和含量比较
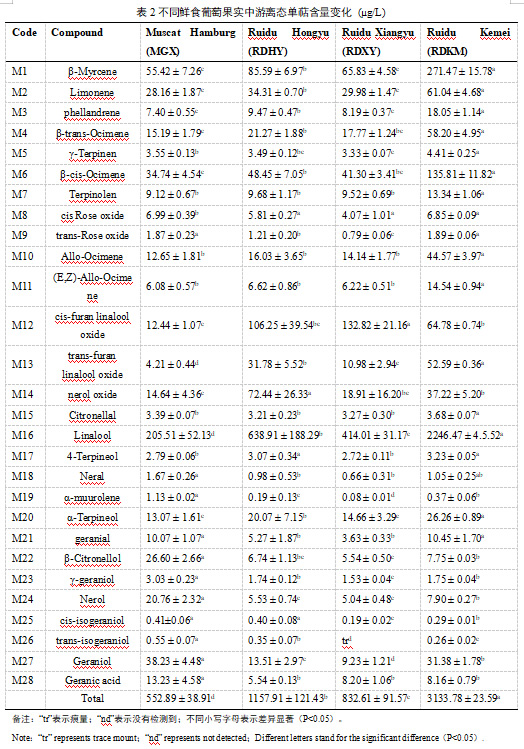
在本研究中,对四个不同葡萄品种中27种游离或糖苷结合形式的单萜(表2和表3)。不同品种单萜的相对丰度差异显著。在四个品种中,‘RDKM’的总游离单萜含量最高(3133.78±23.59 g/L),其次是‘RDHY’( 1157.91±121.43 g/L),‘ RDXY’(832.61±91.57 g/L)和‘MGX’(552.89±38.91 g/L)。在游离单萜类化合物中,芳樟醇(M16)是含量最高的化合物,约占总含量的37.17 % —71.69 %(表2)。除了芳樟醇,β-月桂烯(M1)、香叶醇(M27)、β-顺式罗勒烯(M6)、柠檬烯(M2)和β-香茅醇(M22)也是‘MGX’中的主要单萜;而对于‘RDHY’、‘RDXY’和‘RDKM’,顺式呋喃芳樟醇氧化物(M12)、β-月桂烯(M1)、柠檬烯(M2)、β-顺式罗勒烯(M6)和橙花醇氧化物(M14)的含量相对较高。
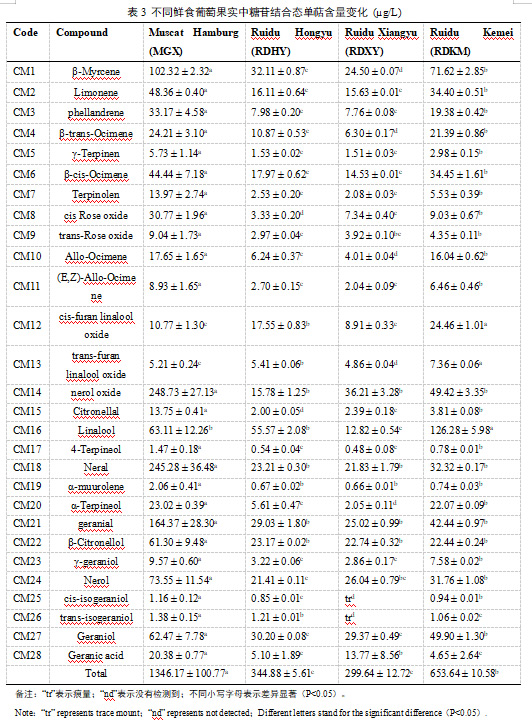
糖苷结合态的单萜表现出与游离形式不同的积累模式(表3)。在四个葡萄品种中,‘MGX’的总糖结合态单萜含量最高(1346.17±100.77g/L),其次是‘RDKM’( 653.64±10.58g/L),其次是‘RDHY’( 344.88±5.61g/L)和‘RDXY’(299.64±12.72)。在‘MGX’葡萄中,糖苷结合态的橙花醇氧化物(CM14)、橙花醛(CM18)、香叶醛(CM21)、β-月桂烯(CM1)、芳樟醇(CM16)被检测为主要的结合态单萜。对于‘RDHY’和‘RDKM’样品,芳樟醇(CM16)、β-月桂烯(CM1)、香叶醇(CM27)、香叶醛(CM21)、β-香茅醇(CM22)和橙花醇氧化物(CM14)是不同品种中的优势化合物。而在‘RDXY’葡萄中,最丰富的糖苷态化合物是橙花醇氧化物(CM14,36.21±3.28g/L),其次是香叶醇(CM27),橙花醇(CM24),香叶醛(CM21)和β-月桂烯(CM1)。
关于不同形态单萜含量的比较,本研究发现除了顺式呋喃芳樟醇氧化葡萄糖苷和4-松油醇葡萄糖苷外,其他26种糖苷键结合的化合物在‘MGX’葡萄中的含量均高于其游离形式(表2和表3),并且糖苷键合单萜的总含量高于游离形式的总含量。而‘RDHY’、‘RDXY’和‘RDKM’葡萄的总糖结合单萜含量低于游离单萜含量,且大多数单萜以游离形式存在。
2.3 低温贮藏过程中单萜化合物变化
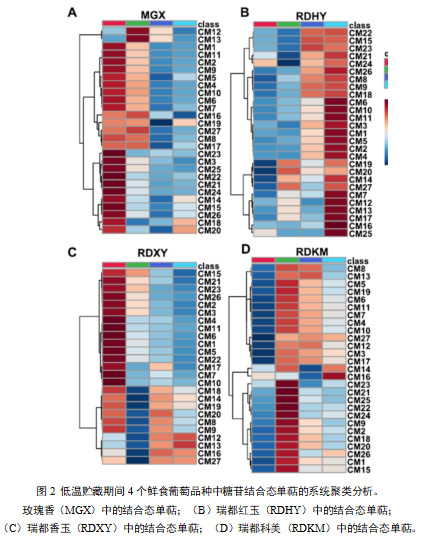
单萜的总含量(游离形式和糖苷键合形式)受到低温贮藏的显著影响(图1和图2)。在低温贮藏期间,所有四种栽培葡萄中游离单萜的总含量均表现出显著下降。在‘MGX’、‘RDHY’和‘RDXY’葡萄中,单萜含量在最初的15天内下降,而在‘RDKM’葡萄中,总游离单萜含量在15天贮藏期间首先上升,然后在随后的贮藏时间内下降。而总结合态单萜呈现不同变化趋势。在‘MGX’和‘RDXY’样品中,总结合单萜浓度在低温贮藏期间下降。但在“RDHY”和“RDKM”样品中,结合单萜的总含量在低温储存期间增加。贮藏期间结合单萜占总单萜的比例总体呈上升趋势。
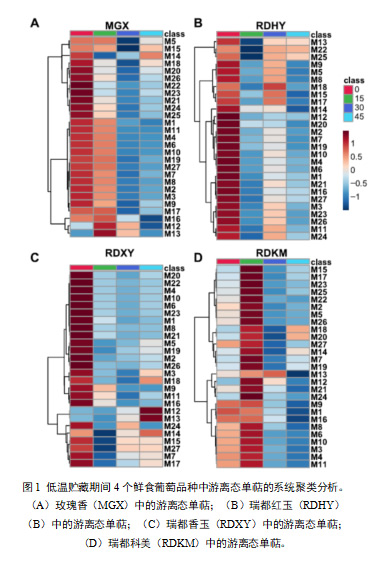
聚类分析结果显示,‘MGX’果实在低温贮藏期间(图1a),这些游离单萜表现出三种主要变化模式。第一种变化模式中的游离单萜包括M5(γ-萜品烯)、M15(香茅醛)、M18(橙花醛)、M20(香叶醛)、M21(β-香茅醇)、M22(γ-香叶醇)和M25(反式异香叶醇)。这些化合物在低温贮藏的前15天下降,在贮藏后30天达到最低水平,但在贮藏后期又上升。在第二种演化模式中,低温贮藏至后期,游离单萜含量逐渐下降,包括M1(β-月桂烯)、M2(柠檬烯)、M3(水芹烯)、M4(β-反式罗勒烯)、M6(β-顺式罗勒烯)、M7(萜品油烯)、M8(顺式氧化玫瑰)、M9(反式氧化玫瑰)、M10(别-罗勒烯)、M19(α-松油醇)和M27(香叶酸)。在第一个15天储存期间,浓度增加后期游离单萜(M12、M13和M16)的下降被聚类为第三种进化模式。
在‘RDHY’中,大多数27种游离单萜烯的浓度在储存开始时(15天)下降,随后在储存后30天上升,然后在储存后45天再次下降(图1b)。聚类分析表明,这些化合物在低温贮藏过程中有两种主要的变化模式。第一组包括M5 (γ-萜品烯)、M8(顺式玫瑰氧化物)、M9(反式玫瑰氧化物)、M15(香茅醛)、M17(4-松油醇)、M18(橙花醛)、M13(反式呋喃芳樟醇氧化物)、M22(γ-香叶醇)和M25(反式-异香叶醇),其在最初15天的贮藏期间显示出浓度的快速下降,然后在贮藏后30天上升。第二类的变化趋势与第一类相似。不同的是,这类化合物的含量在贮藏后30天内略有增加。这类化合物包括M2(柠檬烯)、M3(水芹烯)、M4(β-反式罗勒烯)、M6(β-顺式罗勒烯)、M7(萜品油烯)、M10(别罗勒烯)、M19(α-萜品醇)、M16(芳樟醇)、M21(β-香茅醇)、M23(橙花醇)、M25(反式异香叶醇)和M27(香叶酸)。
在低温贮藏过程中,单萜在‘RDXY’葡萄果实也呈现两种变化模式(图1C)。第一种进化模式中的游离单萜包括M1(β-月桂烯)、M2(柠檬烯)、M4(β-反式-罗勒烯)、M6(β-顺式-罗勒烯)、M8(顺式玫瑰氧化物)、M9(反式玫瑰氧化物)、M10(别-罗勒烯)、M11 ((E,Z)-别-罗勒烯)、M16(芳樟醇)、M20(香叶醛)、M21(β-香茅醇)和M22(γ-香叶醇)。这些单萜类物质的含量一直下降到贮藏后期。在第二种进化模式中,化合物包括M7(松油醇)、M12(顺式呋喃芳樟醇氧化物)、M13(反式呋喃芳樟醇氧化物)、M14(橙花醇氧化物)、M15(香茅醛)、M17 (4-松油醇)和M27(香叶酸)在低温贮藏前15天显著下降,但在低温贮藏后30天又有所上升。
与上述三个品种不同的是,在‘RDKM’样品中,大多数27种游离单萜含量在贮藏前15天显著增加,然后在第30天显著下降(图1d)。在这些化合物上观察到两种主要的变化模式。在第一种演化模式中,单萜类化合物包括M1(β-月桂烯)、M3(水芹烯)、M4(β-反罗勒烯)、M6(β-顺罗勒烯)、M8(反式氧化玫瑰)、M9(反式氧化玫瑰)、M12(顺式呋喃氧化芳樟醇)、M13(反式呋喃氧化芳樟醇)、M16(芳樟醇)、M21(β-香茅醇)和M24(顺-异香叶醇),这些单萜类化合物的含量从贮藏15天开始持续下降,到45天后达到最低水平第二种变化模式中,单萜类物质含量在贮藏后45天内略有增加,包括M2(柠檬烯)、M5(γ-松油烯)、M7(松油烯)、M14(氧化橙花醇)、M15(香茅醛)、M17(4-松油醇)、M22(γ-香叶醇)、M23(橙花醇)、M25(反式异香叶醇)、M26(香叶醇)和M27(香叶酸)。
低温贮藏期间,四个葡萄品种中的结合态单萜与游离态单萜表现出不同的演化模式。在‘MGX’葡萄中,27种结合态单萜在低温贮藏期间表现出两种主要的演变模式(图 2a)。在第一种进化模式中,结合的单萜包括CM8(顺式玫瑰氧化物)、CM12(顺式呋喃芳樟醇氧化物)、CM13(反式呋喃芳樟醇氧化物)、CM16(芳樟醇)、CM17(4-松油醇)、CM19(α-松油醇)和CM 27(香叶酸),其含量在低温贮藏的前15天略有增加,然后在贮藏后30天降至最低。在第二种演化模式中,结合态单萜类化合物含量在整个低温贮藏过程中逐渐下降,包括CM1(β-月桂烯)、CM2(柠檬烯)、CM3(水芹烯)、CM4(β-反式罗勒烯)、CM5(γ-萜品烯)、CM6(β-顺式罗勒烯)、CM7(萜品油烯)、CM9(反式氧化玫瑰酯)、CM10(异罗勒烯)、CM15(香茅醛)、CM21(β-香茅醇)、CM22(γ-香叶醇)、M24(顺式异香叶醇)。在“RDHY”样品中27种结合态单萜含量中也观察到了两种主要变化趋势(图2b)。在这27种化合物中,CM1(β-月桂烯)、CM2(柠檬烯)、CM3(水芹烯)、CM4(β-反式罗勒烯)、CM5(γ-萜品烯)、CM6(β-顺式罗勒烯)、CM7(萜品烯)、CM8(顺式氧化玫瑰)、CM9(反式氧化玫瑰),而其他10种结合态单萜包括CM7(松油醇)、CM12(顺式呋喃氧化芳樟醇)、CM13(反式呋喃氧化芳樟醇)、CM14(氧化橙花醇)、CM16(芳樟醇)、CM17(4-松油醇)、CM19(α-松油醇)、CM20(香叶醛)、CM25(反式-异香叶醇)和CM 27(香叶酸)遵循第二种变化模式,它们的含量在最初的15天贮藏期间略有增加,然后在贮藏后30天下降,但在贮藏后期再次急剧增加。
在‘RDXY’葡萄中,研究了低温贮藏期间27种结合单萜浓度的三种主要演变模式。在第一个实验中,单萜类化合物的含量从贮藏初期开始持续下降,到贮藏45天后降至最低水平,包括CM1(β-月桂烯)、CM2(柠檬烯)、CM3(水芹烯)、CM4(β-反式罗勒烯)、CM5(γ-萜品烯)、CM6(β-顺式罗勒烯)、CM10(别罗勒烯);第二种变化模式中的结合态单萜包括CM12(顺式呋喃氧化芳樟醇)、CM13(反式呋喃氧化芳樟醇)、CM16(芳樟醇)和CM 27(香叶酸)。这些化合物在低温贮藏的前15天内降至最低水平,然后在贮藏后的30天内增加,此后持续增加或在贮藏后期保持在类似水平。与第二种演化模式不同,第三种模式中的单萜在贮藏后期浓度呈下降趋势。这些化合物包括CM8(顺式氧化玫瑰)、CM9(反式氧化玫瑰)、CM14(氧化橙花醇)、CM17(4-松油醇)、CM18(香叶醛)、CM19(α-松油醇)和CM20(香叶醛)。在‘RDKM’样品中的27种结合单萜烯中,除了CM 14(氧化橙花醇)和CM16(芳樟醇)之外,大多数都呈现出相似的演化模式(图 2d),其含量在贮藏30天后下降,但在贮藏45天后又上升。其他指标在贮藏的前15天呈上升趋势,之后逐渐下降直至贮藏结束。
2.4 低温贮藏过程中转录组分析
为了进一步探索上述鲜食葡萄低温贮藏期间单萜差异的分子基础,进行了RNA-seq。基于FPKM值计算基因表达水平(绘制的每百万个阅读的外显子模型的每千个碱基的片段数),然后进行皮尔逊相关和PCA分析。所有样品都显示出一定程度的相关性。系统聚类分析表明,所有三个品种的新鲜葡萄样品与低温贮藏后的样品之间的基因表达差异明显,不同品种之间也存在显著差异。类似于聚类的结果,不同贮藏阶段和不同品种之间显著分离,表明低温储存期间许多基因的表达水平发生变化。样本之间的DEGs用log2(倍数变化)≥1和FDR<0.05进行筛选。
通过GO功能富集和KEGG进一步分析DEGs的功能。通过比较不同贮藏阶段的样品得出的DEGs在 “代谢过程”、“细胞过程”和“生物调节”等生物过程方面显著富集;在 “细胞”、“细胞部分”和“细胞器”等细胞水平富集;以及“结合”和“催化活性”等分子功能层面富集。KEGG分析结果表明,在低温贮藏期间,大多数DEGs在“次生代谢物的生物合成”、“代谢途径”、“类苯丙酸生物合成”、“植物激素信号转导”和“植物-病原体相互作用”等途径中富集。
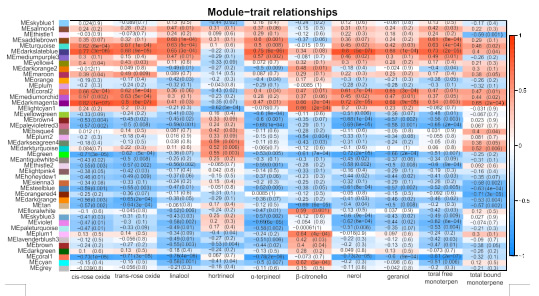
为了对共表达模块进行分类并鉴定与低温贮藏期间单萜含量变化相关的关键候选基因,使用所有检测到的转录物和10个代表性的游离单萜进行了WGCNA分析(图3)。具有相同表达模式的基因被分组到相同的模块中,形成聚类树状图。在这项研究中,从RNA-Seq数据中揭示了总共43个模块(图3),其中青绿色模块包含最多的基因(3986个基因),而紫红色模块包含最少的基因(64个基因)。模块与单萜含量之间的相关系数变化很大,从0.81到0.82(图3)。发现其中三个模块(darkslateblue、darkmagenta和coral1模块)与大多数单萜的含量最显著相关,表明这些模块中的基因与单萜含量显著相关。除hortrineol外,darkslateblue和darkmagenta模块中的基因与单萜含量呈正相关,而coral1模块中的基因与所有单萜含量呈负相关。进一步分析了三个模块中固有基因的生物学功能。如结果所示,在模块darkslateblue中显著富集5个KEGG途径,包括不饱和脂肪酸的生物合成脂肪酸、碳代谢、氨基酸的生物合成等。模块coral1中富集了11个KEGG途径,包括精氨酸和脯氨酸代谢、苯丙氨酸代谢、植物激素信号转导等。值得注意的是,2个KEGG途径在darkmagenta模块中显著富集,包括萜类骨架生物合成(ko00900),表明该模块在参与低温贮藏期间单萜化合物变化中的重要性。萜类骨架生物合成途径注释到14DEGs(Vitvi02g00420、Vitvi03g00374、Vitvi06g00683、Vitvi07g01408、Vitvi07g01917、Vit vi07g04151、Vitvi11g01303、Vitvi12g02534、Vitvi12g02536、Vitvi14g00258、Vitvi17g00241)。

2.5 qRT-PCR分析
此外, qRT-PCR结果(图4)显示了与RNA-seq分析相似的变化模式,表明转录组数据的可靠性。单萜生物合成的MEP途径中的大多数基因在低温贮藏期间表达量下调,包括3个1-脱氧-D-木糖-5-磷酸合酶(DXS)基因(Vitvi04g00438,Vitvi05g00372和Vitvi07g01408)、2-C-甲基-D-赤藓醇4-磷酸胞苷酰转移酶(MCT)基因(Vitvi12g02141)、4-(胞苷5′-二磷酸)-2-C-甲基-D-赤藓醇激酶(CMK)基因(Vitvi06g01125)、2-甲基-D-赤藓醇2,4-环二磷酸合酶(MDS)基因(Vitvi06g00286)、4-羟基-3-甲基丁-2-烯二磷酸还原酶(HDR)基因(Vitvi03g00374)、香叶基二磷酸合酶(GPPs)基因(Vitvi19g00506)、4个萜类合成酶(TPS)基因。而MVA途径中的基因包括2个3-羟基-3-甲基葡萄糖- taryl-CoA合酶(HMGS)基因(Vitvi02g00420和Vitvi17g00241)、2个3-羟基-3-甲基戊二酰-CoA还原酶(HMGR)基因(Vitvi03g00262和Vitvi04g01740)、1个牻牛儿基牻牛儿基二磷酸合酶(GGPPs)基因(Vitvi18g00922)等,表达量呈现上升趋势。
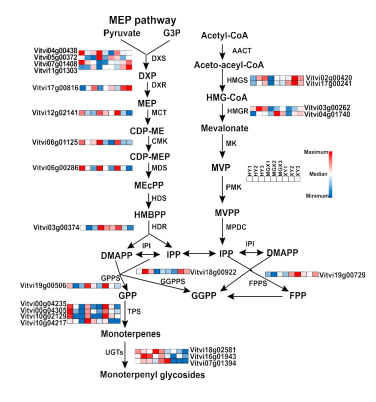
图4 3个葡萄品种单萜生物合成途径在不同贮藏阶段的差异表达分析。蓝色到红色表明基因表达逐渐增加。MEP途径:DXS,1-脱氧-D-木酮糖-5-磷酸合酶;1-脱氧木酮糖-5-磷酸还原异构酶;MCT,4-二磷酸缩水甘油基-2-C-甲基-D-赤藓糖醇合酶;CMK,4-二磷酸胞酰-2-甲基-D-赤藓糖醇激酶;MDS,2-C-甲基-D-赤藓糖醇2,4-环二磷酸合酶;1-羟基-2-甲基-2-(E)-丁烯基4-二磷酸合酶;1-羟基-2-甲基-2-(E)-丁烯基4-二磷酸HDR还原酶;IPI,异戊烯二磷酸异构酶;GPS,牻牛儿基二磷酸合酶;UGT,UDP-糖基转移酶;牻牛儿牻牛儿二磷酸合酶;TPS,萜烯合酶;MVA途径:ACAT,乙酰辅酶a乙酰转移酶;HMGS,3-羟基-3-甲基戊二酰辅酶a合酶;HMGR、3-羟基-3-甲基戊二酰辅酶a还原酶;MK、甲羟戊酸激酶;PMK磷酸甲羟戊酸激酶;MPDC甲羟戊酸二磷酸脱羧酶;法呢基二磷酸合酶;MVA;甲瓦龙酸;甲基赤藓糖醇磷酸酯,IPP;异戊烯基二磷酸酯;二甲基烯丙基二磷酸酯;二磷酸法呢基FPP;GPP,二磷酸香叶酯。