酿酒葡萄品种改良岗位
摘要:尿苷二磷酸糖基转移酶(UGTs)参与糖基化修饰过程,在植物生长发育、次生代谢和逆境响应中发挥重要作用。本文对UGT家族进行了系统的全基因组分析,以探讨其在葡萄中的重要功能。共鉴定出181个VvUGT,分布在18条染色体上。基于系统发育分析,这些UGT基因被聚为17个主要类群(A-P, R),具有保守的基因结构和一致的基序。对VvUGT基因的共线性分析表明,片段重复和串联重复事件对VvUGT基因家族的进化有显著影响。这些研究结果为进一步开展VvUGTs的功能解剖和酿酒葡萄的遗传改良奠定了重要基础。
糖基化是所有生物体内的重要修饰。糖基转移酶(GTs)在各种代谢过程中负责糖基化修饰,普遍存在于所有生物体中。其中,家族-1糖基转移酶,也称为UDP糖基转移酶(UGTs),是植物中最常见的糖基转移酶。UGT能够催化糖基转移到植物中的特定受体,包括植物激素、次生代谢物和外源物。目前,随着植物基因组测序的完成,UGT在各种植物中得到了广泛的研究。在拟南芥中发现了100多个UGT基因。在其他植物物种中也发现了更多的UGT成员,如玉米(147)、小麦(179)、柑橘(145)、苹果(214)和桃子(168)。
葡萄(Vitis vinifera L.)是世界上非常重要的水果作物,其果实含有特殊的风味和丰富的营养成分。它可以新鲜或干燥食用,也可以加工成各种产品,如果酱、果汁、葡萄酒、醋和葡萄籽油。在这项研究中,我们对葡萄UGT基因家族进行了广泛的生物信息学分析,并探索了VvUGT的潜在功能。基于葡萄基因组共鉴定出181个成员。进行了系统发育分析、染色体定位和共线性分析。该研究将为进一步研究葡萄UGT基因的功能特性提供重要依据。
1 材料与方法
1.1 葡萄UGT基因家族成员鉴定
从Phytozome V13下载葡萄蛋白序列(https://phtozome-next.jgi.doe.gov/)。拟南芥UGT序列来源于TAIR (https://www.arabidopsis.org/)。本研究采用两种方法对VvUGT基因进行鉴定。首先,以拟南芥UGT序列为查询对象,在葡萄蛋白数据库中进行blastp检索。其次,从Pfam网站 (http://pfam.xfam.org/)中检索UGT结构域(PF00201)的the Hidden Markov Model (HMM)。利用HMM文件和hmmsearch软件从葡萄蛋白数据中鉴定出推测的VvUGT基因,e值阈值为1.0e-5。用SMART对两种方法产生的候选成员进行筛选,去除不含完整PSPG基序的蛋白质。使用ExPASy Online程序(http://web.expasy.org/protparam/)分析了VvUGT的基本特性。
1.2 系统发育进化分析
将葡萄、拟南芥、番茄、山柳兰和玉米的UGTs蛋白序列利用MUSCLE进行比对,并利用MEGA 11构建了1000个bootstrap重复的邻居连接(NJ)系统发育树。根据葡萄GFF3注释文件中的内含子-外显子数据,利用TBtools软件绘制了VvUGT的基因结构。使用MEME (https://meme-suite.org/meme/tools/meme)鉴定VvUGT蛋白的保守基序,每个序列最多10个基序。
1.3 VvUGT染色体定位及共线性分析
利用TBtools软件根据葡萄基因组的定位信息绘制了VvUGT的染色体定位图谱。使用带有默认参数的多个共线扫描工具(MCScanX)搜索葡萄基因组中UGT基因的串联重复和片段重复事件。利用MCScanX对葡萄和拟南芥的UGT进行了共线性关系分析。
2 结果与分析
2.1葡萄UGT基因鉴定和染色体位置信息
在葡萄基因组中共鉴定出181个UGT基因。这些UGT根据其染色体位置被命名为VvUGT1至VvUGT181。除10个UGT成员外,它们在18条葡萄染色体上分布不均匀如图1所示。其中,5号染色体含UGT基因最多(26个成员),其次是18号染色体(23个成员),12号染色体(19个成员)。只有一个成员被定位到9号染色体上。值得注意的是,在Chr3、Chr5、Chr6和Chr17等18条染色体上观察到许多簇。

2.2 葡萄UGT基因家族成员系统发育、基因结构及保守基序分析
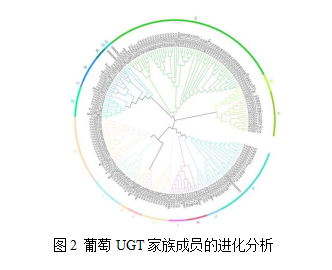
以葡萄、拟南芥、番茄、山柳兰和玉米的UGT蛋白序列为基础,构建了VvUGT基因家族的系统发育树。由图2可知,系统发育树显示,181个VvUGT可分为17组(A-P, R)如图5-2所示。E组包含最多的VvUGT成员(56个),其次是P组(24个)。O组是最小的(只有一个VvUGT成员)。Q组只在玉米中出现,说明可能是单子叶特异性组。
2.3 种间和种内共线性分析
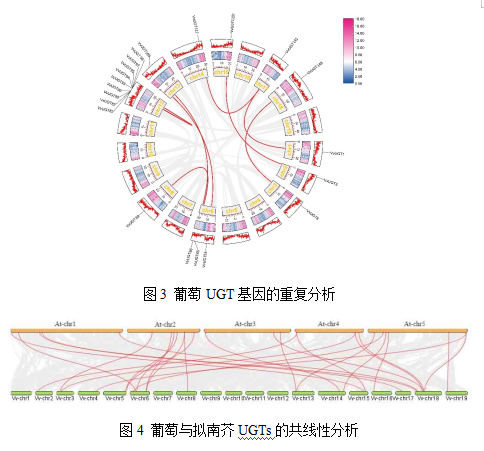
为了探究VvUGT基因家族的扩增与进化机制,我们分析了葡萄基因组中的基因重复事件如图3所示。本研究利用MCScanX软件,基于氨基酸序列同源性对葡萄基因组进行比较。共鉴定出15对片段重复基因对,包括VvUGT85/93、VvUGT85/98、VvUGT149/8、VvUGT1/41、VvUGT89/99、VvUGT191/3、VvUGT117/141、VvUGT65/69、VvUGT90/95、VvUGT109/66、VvUGT82/93、VvUGT82/97、VvUGT104/53、VvUGT85/94和VvUGT86/94。此外,染色体内重复分析显示,159个VvUGT基因来自串联重复事件,导致基因组中的同源基因簇。这些结果表明,片段复制和串联复制在VvUGT家族的扩展中发挥了重要作用。
为了进一步探索VvUGT基因家族的进化过程,我们对拟南芥和葡萄进行了共线性分析如图4所示。拟南芥和葡萄基因组共线性UGT基因对共有27对,其中24个基因来自拟南芥,27个基因来自葡萄。这些VvUGT位于13条染色体上(chr2、3、4、5、6、8、13、14、15、16、17、18和19),其中6号和18号染色体共线VvUGT基因最多。VvUGT基因在染色体上的不均匀分布表明在葡萄的进化过程中存在遗传变异。
3 讨论与结论
UGT参与植物次生代谢产物的糖基化和修饰,在植物生长发育和逆境响应中发挥重要作用。葡萄是世界上重要的水果作物之一,可用于鲜食还可以制作果汁和葡萄干,它也是葡萄酒行业的重要原料。同时UGT基因参与并影响葡萄单宁的合成与代谢,因此,本研究对影响葡萄单宁合成与代谢的UGT基因家族进行了系统分析,共鉴定出181个VvUGT基因。与柔毛淫羊藿 (339)、紫花苜蓿(409)和花生(267)相比, 葡萄UGT的数量少,但多于玉米(147)、桃子(168)、石榴 (120)和茶(178)。根据系统发育分析,181个VvUGT被划分为(A-P、R) 17个组,其中E组中VvUGT成员最多(56个),占30.9%。值得注意的是,在Q组中没有VvUGT成员,但在玉米中发现了VvUGT成员。在小麦中也发现了类似的现象,推测Q群可能只存在于单子叶植物中。
在植物进化过程中,基因复制促进了基因家族的扩展。在我们的研究中,在葡萄基因组中鉴定了15个片段重复和103个串联重复,说明串联复制是VvUGT家族扩增的主要动力。这可能就是为什么VvUGT基因在染色体上以基因簇的形式不均匀分布的原因。另外拟南芥和葡萄UGT家族共线性分析显示,E组的同源对最多,其次是P组。A、B、C和F组的共线性对最少(只有1对)。这些结果表明,E组成员可能为UGT基因家族的起源和进化提供了更多线索。此外,我们发现VvUGT150在拟南芥中有3个同源基因,说明葡萄基因组进化过程中存在基因丢失事件。UGT基因的进化历史与其功能密不可分,已有研究表明VvUGT基因参与并影响葡萄单宁的合成与代谢,因此,研究VvUGT基因的进化历史及结构对探究影响葡萄单宁合成与代谢的UGT基因家族具有重要意义。