豫东综合试验站
张柯 娄玉穗 尚泓泉 吕中伟
经过长期的栽培、驯化、杂交育种、营养系选种和引种交流,葡萄形成了复杂基因渗入的欧美杂种、山欧杂种等杂种群,葡萄的表型及基因型都呈现出极丰富的遗传多样性。由于葡萄的无性繁殖,容易扦插扩繁,跨地域引种和资源交换频繁,导致命名重复、同名异物和品种名丢失现象普遍发生,造成葡萄种类数目不清、种名混乱。传统的形态学鉴定法难以有效区分源自少数育种骨干亲本的众多新品种,给葡萄资源利用造成很大的困难。利用分子标记技术可有效鉴定种质资源,分析品种间的遗传多样性。SLAF-seq具有通量高、成本低、准确性高的优点,可广泛应用于亲缘关系和遗传进化分析、遗传图谱构建等研究。本研究选取了前人未进行简化基因组测序研究的23份鲜食葡萄种质,包括生产中的主栽品种、国内外新引进品种和部分系谱不明的优良种质,利用SLAF-seq技术对其进行测序,在全基因组范围内鉴定葡萄的多态性SNP标记,基于SNP进行遗传关系分析,以探明遗传背景不清楚的葡萄种质的亲缘关系,为葡萄种质资源的科学利用和种质创新提供理论参考。
1 材料和方法
1.1 材料
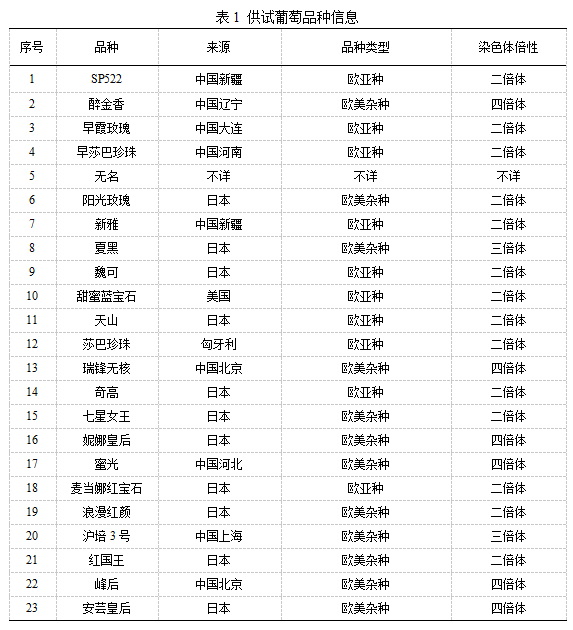
选取生产中的主栽品种、国内外新引进品种和部分遗传背景不明的优良种质共23份,包括10份欧亚种、12份欧美杂种和1份品种类型不清楚的种质(表1)。所有试验材料于2020年4月采自国家葡萄产业技术体系豫东综合试验站核心示范园。园区内葡萄株行距1.5m× 3m,避雨栽培模式,高宽垂架,常规生产管理。
1.2 方法
于萌芽期取各材料的幼嫩叶片,锡箔纸包裹,做好标记,于液氮中冻存带回实验室,用改良CTAB法提取DNA,分别用1.0%琼脂糖凝胶电泳法和NanoDrop1 000C分光光度计法检测DNA的完整性和浓度。将合格的DNA用超纯水将浓度稀释至50 ng/μL,于-20 ℃冰箱保存,用于后续的测序、构建文库。
SLAF测序:选取已测序完成的葡萄(Vitis vinifera L.)基因组为参考基因组,参考基因组约为416 Mb,GC含量为39.08%,组装出的基因组为486 Mb,GC含量为34.55%。利用限制性内切酶切组合Rsa I + Hae III进行酶切,质量检测合格后测序、建库、质检。为保证测序质量,采用读长126 bp×2分析数据,用Q30评估测序质量。以水稻‘日本晴’(Oryza sativa ssp. Japonica)基因组数据为对照,对测序文库进行评估。用Illumina Hi Seq 2500进行双端测序,利用SOAP软件进行参考基因组比对。‘日本晴’基因组大小为 374.30 Mb。利用Dual-index识别测序的raw reads进行质控,评估测序质量。统计‘日本晴’与其参考基因组的双端比对效率、酶切效率、测序reads插入片段分布,评估测序文库。利用BWA软件将clean reads和SLAF标签分别比对定位到参考基因组上,统计比对率、各样品测序深度、SLAF标签及多态性SLAF标签在不同染色体上的分布情况。将两种软件GATK 和SAM tools开发的SNP标记交集,作为群体SNP标记数据集。对开发的SNP分子标记进行过滤,过滤标准为MAF(Minor allele frequency, 次要等位基因频率)>0.05,完整度>0.8,通过统计软件Admixture、Eigensoft和MEGA 5,利用高质量的SNP进行亲缘关系分析。
2 结果与分析
2.1 酶切与方案评估
选用Rsa I和Hae III限制性内切酶组合对葡萄的参考基因组进行酶切位点分析,预测可得到101 869个片段长度在314-464 bp的SLAF标签。以‘日本晴’的reads作为对照评价实验的成功性。‘日本晴’测序reads的双端比对效率为96.84%,对比效率高;Rsa I+Hae III正常的酶切效率为86.12%,酶切效率好,表明本实验酶切方案可行。
2.2 质量评估
对照‘日本晴’获得1.21Mb reads的数据量,从23个葡萄品种共获得127.95 Mb clean reads,25.96Gb Clean Data。各样品的reads数分布在983 965 bp~10 110 544 bp范围内,平均5 563 097.96bp。其中,reads数据量最大的葡萄品种是‘醉金香’,最小的品种是‘早霞玫瑰’。各样品的GC含量分布范围在37.79%~41.39%,平均GC含量为39.89%,对照的平均GC含量为43.35%。各样品的Q30分布范围在89.83%~94.99%,平均值为93.93%,对照的Q30为93.78%。Q30值很高,表明本次测序的碱基错误率极低,测序质量很高,测序结果是可靠的。
2.3 SNP开发
对23份葡萄样品进行简化基因组测序,平均测序深度为15.58x,每个样品平均开发283 515个SLAF标签,共获得715 776个SLAF标签,其中多态性SLAF标签共有505 092个共得到4 026 432个群体SNP,每个样品的SNP数目从1 191 974至2 626 279不等,平均完整度为29.60%~65.23%。SNP 杂合率为6.76%~27.33% 。不同样品,存在一定的差异,表明葡萄基因组杂合度很高。
2.4 遗传关系分析
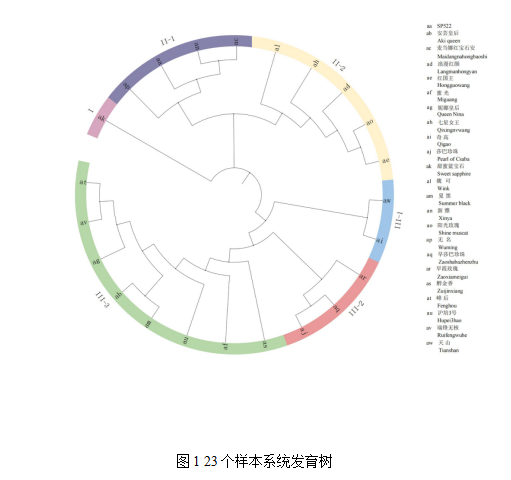
基于456 776个高质量的SNP,对23份葡萄种质进行群体结构分析,构建进化树 (图1),从基因组水平揭示种质资源的遗传分化关系。所有种质按遗传距离远近可聚为3大类。组Ⅰ只有1份葡萄种质,‘甜蜜蓝宝石’,为美国育成的大粒无核葡萄品种,其亲本不详。‘甜蜜蓝宝石’与其余22份种质的亲缘关系较远,虽未找到与‘甜蜜蓝宝石’相近的种质,但至少说明‘甜蜜蓝宝石’与骨干育种亲本‘巨峰’、‘红地球’、‘玫瑰香’和‘莎巴珍珠’遗传距离较远。组Ⅱ包括9个葡萄种质,可分为2个亚组,均为二倍体。Ⅱ-1中有4个种质,同属欧亚种。‘新雅’为‘红地球’和‘里扎马特’的衍生后代,‘SP522’为‘里扎马特’的子代,‘麦当娜红宝石’和‘无名’(丢失品种名)的亲本不详,聚类分析结果表明两者与‘新雅’、‘SP522’的遗传距离较近,结合生长和果实特性,推断‘无名’可能有‘红地球’的血统,‘麦当娜红宝石’可能有‘里扎马特’的血统。Ⅱ-2中有5个种质,以欧美杂种为主,只有‘魏可’为欧亚种。‘魏可’为‘甲斐路’杂交后代;‘阳光玫瑰’为欧美杂种,其父本‘白南’有‘甲斐路’的血统,其母本有司特本的血统,司特本为美洲葡萄和欧亚葡萄的杂交种。分属不同种群的‘魏可’和‘阳光玫瑰’可聚为一个亚组,其原因是都有‘甲斐路’的血缘血统。‘七星女王’、‘浪漫红颜’、‘红国王’为‘阳光玫瑰’的杂交后代,同属欧美杂种。组Ⅲ最为复杂,包括13个种质,既有二倍体欧亚种、三倍体欧美种,又有四倍体欧美种。组Ⅲ又可分成3个亚组。Ⅲ-1包括‘天山’和‘奇高’2个种质,同属二倍体欧亚种,均有‘白罗莎里奥’血统,而‘白罗莎里奥’为‘罗沙克’和‘白玫瑰香’的杂交后代。Ⅲ-2有3个种质,均为二倍体欧亚种,具有玫瑰香味。‘早霞玫瑰’为‘白玫瑰香’和‘秋黑’的杂交后代,‘莎巴珍珠’为‘匈牙利玫瑰’和‘奥托涅玫瑰’的杂交后代,‘早莎巴珍珠’为‘莎巴珍珠’的早熟芽变。Ⅲ-3包括8个种质,同属欧美种,均为‘巨峰’的衍生后代。‘巨峰’的父本为‘罗沙克’的四倍体芽变。因为共同拥有‘罗沙克’的血统,二倍体欧亚种‘天山’、‘奇高’和欧美种‘巨峰’的衍生后代和聚为一个组。四倍体欧美种‘醉金香’和二倍体欧亚种‘早霞玫瑰’均为‘白玫瑰香’的衍生后代,因此聚在了一个组。可以看出,利用SLAF-seq构建进化树从基因水平上鉴定葡萄种质的遗传关系是可靠的。
3 结论与讨论
葡萄种质的基因型和遗传关系的确立是分子育种的基础。本研究以前人未进行SLAF测序分析的23份鲜食葡萄种质为材料,利用SLAF-seq技术在全基因组范围筛选456 776个高质量SNP标记,分析了23份葡萄种质的系谱、遗传背景和亲缘关系。结果表明,姊妹系被聚在一类,如‘浪漫红颜’和‘红国王’;子代和亲本聚在一起,如‘阳光玫瑰’和‘七星女王’、‘浪漫红颜’、‘红国王’;‘巨峰’系多倍体品种聚类到到欧美杂交种中。此外,分属不同种群的葡萄种质聚类在了一起,如欧美种‘阳光玫瑰’及其衍生品种与欧亚种‘魏可’聚类到一组,而欧亚种‘天山’、‘奇高’、‘早霞玫瑰’与‘巨峰’系聚在了欧美种群。分属不同种群的葡萄种质聚类在一组的原因是因为拥有共同的血缘血统。本研究中,欧亚种‘魏可’和欧美种‘阳光玫瑰’均有‘甲斐路’的血缘血统,二倍体欧亚种‘天山’、‘奇高’和欧美种‘巨峰’的衍生后代共同拥有‘罗沙克’的血统,四倍体欧美种‘醉金香’和二倍体欧亚种‘早霞玫瑰’均为‘白玫瑰香’的衍生后代。基于SLAF-seq的群体结构分类和系谱分类的结果具有高度一致性,说明利用SLAF-seq构建遗传进化树从基因水平上鉴定葡萄种质的遗传关系是可靠的。
本研究基于SLAF-seq的群体结构分析,将具有相同遗传背景的葡萄种质聚为了一类,为系谱不明和丢失品种名的葡萄种质找到了遗传距离近的品种,很好地探明其遗传背景,为葡萄种质鉴定、资源保护和高效育种提供了参考依据。