华中西南区栽培岗位
熊榆 许延帅 谭君 杨国顺
团队前期研究结果表明同一品种延后果与正常果实挥发性香气物质含量有较大不同,本研究以‘夏黑无核’、‘户太八号’、‘巨玫瑰’、‘瑞都红玫’延后果作为试验组,正常果为对照组进行转录组测序,通过不同熟期葡萄香气合成途径上关键基因表达差异分析,研究延后果与正常果香气物质差异形成机理。
1.材料与方法
四个品种葡萄正常果与延后果完全成熟果实(E-L 38),共8组样品。
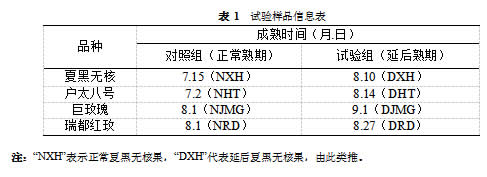
1.1 样品采集与保存试验方法
选取四个品种成熟果实250 g立即用镊子将果皮剥下,去籽,随后将果肉切碎用液氮速冻,并马上将速冻样品转入有标签的小离心管中。所有操作在田间完成后迅速带回实验室放入-80 ℃超低温冰箱中保存。
1.2 RNA提取、文库构建与转录组测序,数据处理
RNA提取以及转录组测序委托北京百迈客生物科技有限公司完成。使用Biomarker Plant Total RNA Isolation Kit(Poly-saccharides&Polyphenolics-rich)试剂盒提取RNA后检测RNA样品纯度浓度以及完整性。待样品检测合格后,进行文库构建,主要流程如下:
(1) 用带有Oligo(dT)的磁珠富集真核生物mRNA;
(2) 加入Fragmentation Buffer将mRNA进行随机打断;
(3) 以mRNA为模板,用六碱基随机引物(random hexamers)合成第一条cDNA链,然后加入缓冲液、dNTPs、RNase H和DNA polymerase I合成第二条cDNA链,利用AMPure XP beads纯化cDNA;
(4) 纯化的双链cDNA再进行末端修复、加A尾并连接测序接头,然后用AMPure XP beads进行片段大小选择;
(5) 最后通过PCR富集得到cDNA文库。
文库构建完成后使用实时荧光定量(Q-PCR)技术对构建文科的有效浓度进行精准定量,以确保文库质量。库检合格后进行样本混合(Pooling),使用第二代测序技术illumina平台进行测序。
数据下机后进行过滤得到Clean Data,与指定参考基因组(https://plants.ensembl.org/Vitis_vinifera/Info/Index)进行序列比对,得到Mapped Data,并对插入片段长度,随机性等文库质量项目进行评估。
数据处理与制图在百迈客云平台(https://international.biocloud.net/)完成。
2. 结果与分析
2.1 测序质量评估
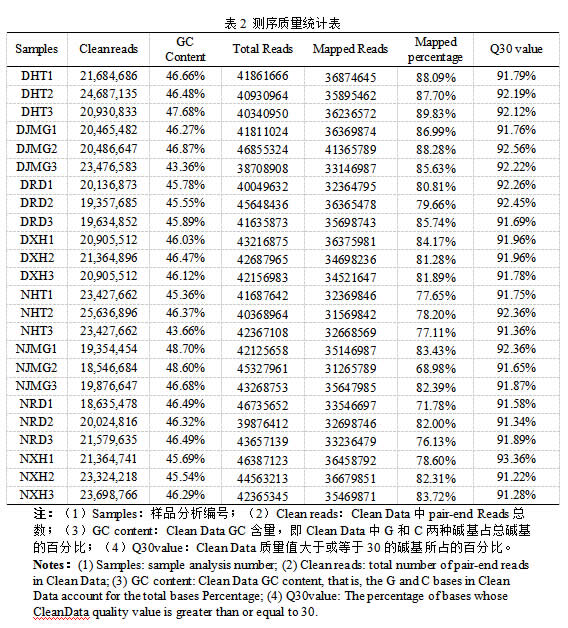
共得到151.47 Gb Clean Data,有80~90% reads对比到参考转录组上,其中比率较高的有DHT、DJMG、DXH。所有样品碱基百分比(Q30)含量均高于91.22%,表明测序数据可靠性良好,可进行下一步分析。
2.2 四个品种延后与正常果差异表达基因分析
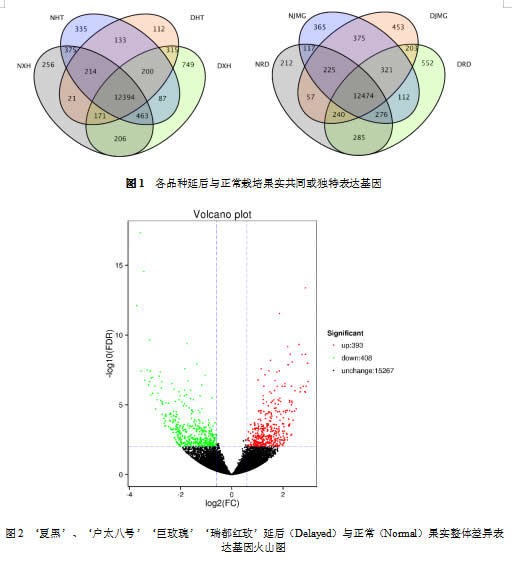
对四个品种延后与正常果之间共同与独特表达基因(DEGs)进行筛选,差异表达阈值设定为1.5。
‘夏黑’延后与正常果共有2929个基因显著上调或下调,其中下调基因1665个,主要有Vitvi14g01808(K10529,α-亚麻酸代谢)、Vitvi13g00471(K13993,HSP20家族蛋白)、Vitvi05g00483(K00276,氨基脲敏感的胺氧化酶)等;上调基因有1264个,包含Vitvi02g00230(K20929,乙二醛/甲基乙二醛氧化酶)、Vitvi07g02514(K14638,肽/组氨酸转运蛋白家族15)、Vitvi07g02517(K08233,聚神经苷醛酯酶)等; ‘户太八号’延后果有2161个基因发生显著上调或下降,为四个品种中最少,其中下调基因961个,有Vitvi13g02014(K13993,HSP20家族蛋白)、Vitvi07g02628(K03937,NADH脱氢酶(泛醌Fe-S蛋白)、Vitvi01g00922(K20929,乙二醛/甲基乙二醛氧化酶)等,上调基因有1200个,其中Vitvi03g00104(K01802,肽基脯氨酰异构酶)、Vitvi12g00261(K00588,咖啡酰-CoA O-甲基转移酶)基因差异表达|Log2FC|值较大;‘巨玫瑰’延后与正常果差异表达基因共有2385个,上调/下调基因分别为1267/1118个,其中下调基因中有Vitvi04g00092(K09419,热激转录因子)、Vitvi14g02043(K02160,乙酰辅酶A羧化酶生物素羧基载体蛋白)、Vitvi18g01063(K20929,乙二醛/甲基乙二醛氧化酶),上调基因主要有Vitvi12g00574(K12742,异戊二烯合酶),Vitvi18g02730(K20858,钙单转运蛋白)、Vitvi19g01726(K14709,锌转运蛋白)等;‘瑞都红玫’延后与正常果差异表达基因有2652个,其中1122个基因出现显著下调,包括Vitvi16g00054(K10775,苯丙氨酸解氨酶),VviSTS35(K00660,查尔酮合酶),VviSTS47b(K13232,二苯乙烯合酶)Vitvi01g00922(K20929,乙二醛/甲基乙二醛氧化酶)等,同时1530个基因出现上调,上调基因有Vitvi16g01082(K19747,BAHD酰基转移酶),Vitvi17g00615(K08910,捕收复合体I叶绿素a/b结合蛋白)等(图1)。
通过对四个品种整体延后与正常果差异表达基因进行统计,发现共有393个基因出现显著上调,408个基因发生下调,其中Vitvi04g00932(K00924,受体蛋白激酶TMK1)、Vitvi05g00953(K01728,果胶酸裂合酶)基因显著上调,而Vitvi02g01446(K13993,HSP20家族蛋白)、Vitvi02g00025(K04079,molecular chaperone HtpG)、Vitvi09g00375(K06215,磷酸吡哆醛酶)等基因出现显著下调。
2.3 ‘夏黑’‘户太八号’‘巨玫瑰’延后与正常果脂氧合酶与氨基酸途径相关基因的转录表达差异分析

脂氧合酶途径与氨基酸途径主要合成果实中的酯类与C6/C9醇醛类物质,其中果香型与酯香型果实的特征挥发性香气物质均由氨基酸途径合成;C6/C9醇醛类物质给葡萄果实带来绿叶味清香,所以又称“绿叶挥发物”(GLVs),它们通常由脂氧合酶途径合成。脂肪氧化酶(LOX)作为脂氧合酶途径第一个酶,能够调节植物信号分子产生,果实成熟,胁迫防御等生理过程。表4.2中延后 ‘巨玫瑰’VviLOXA基因出现显著上调,而其余LOX酶基因出现下调;延后‘户太八号’与‘夏黑’测得大部分LOX酶基因出现显著上调(Vitvi06g00158、Vitvi13g01780、Vitvi05g00472),而Vitvi01g01562基因出现显著下调。过氧化物裂解酶(VviHPL)也是脂氧合酶途径中重要的合成酶,VviHPL与果实内C6/C9醛含量密切相关,研究认为植物叶片细胞中挥发性醛类化合物的合成依赖于HPL酶活性,本试验中延后‘夏黑’‘户太八号’‘巨玫瑰’VviHPL1基因均上调表达。酰基转移酶(VviAAT)与醇脱氢酶(VviADH)也是氨基酸途径中重要的反应酶,VviADH与VviAAT可在脂氧合酶途径与氨基酸途径中依次将醛转化成醇,再将醇催化成相应的酯。本试验在延后‘巨玫瑰’‘户太八号’‘夏黑’中得到了9条ADH基因,其中Vitvi17g00283、Vitvi04g01668、Vitvi04g01669、Vitvi04g01671、Vitvi18g01226基因在三个品种延后果中均出现显著下调,而Vitvi14g01522与Vitvi07g03077则出现上调。延后‘夏黑’得到的ADH酶基因只有Vitvi14g01522出现明显上调。本试验中延后‘夏黑’‘户太八号’‘巨玫瑰’得到的VviHPL1,VviAAT基因均出现显著上调。
2.3 ‘巨玫瑰’’瑞都红玫’延后与正常果萜类物质合成途径相关基因的转录表达差异分析
萜烯类是玫瑰香型葡萄重要的成香物质之一,其中香叶醇,芳樟醇,香茅醇等单萜类化合物为玫瑰香型葡萄带来了浓郁的玫瑰香与花香风味。单萜类物质主要由异戊烯焦磷酸(IPP)与二甲烯丙基焦磷酸(DMAPP)通过异戊烯转移酶的作用下生成,而IPP与DMAPP由甲羟戊酸途径(MVA)途径与2-C-甲基-D-赤藓糖醇-4-磷酸(MEP)途径合成。
MEP途径中共得到2条1-脱氧木酮糖-5-磷酸合成酶(VviDXS)基因与1条1-脱氧-D-木酮糖-5-磷酸还原异构酶(VviDXR)基因差异表达,其中延后‘巨玫瑰’中VviDXS、VviDXR基因显著上调;‘瑞都红玫’中VviDXS基因显著上调,而VviDXR基因出现下调。MVA途径中得到2条乙酰辅酶A酰基转移酶(VviAACT)基因与表达发生显著变化,其中Vitvi10g01337基因在延后‘瑞都红玫’中显著下调但并未在延后‘巨玫瑰’中表达,而Vitvi09g00996在延后‘巨玫瑰’中表达下调,在延后‘瑞都红玫’中没有表达。3-羟基-3-甲基戊二酰辅酶A还原酶(HMGR)是MVA合成途径上重要的限速酶,本试验中2条VviHMGR基因在延后‘巨玫瑰’中出现下调,而在延后‘瑞都红玫’中显著上调。
两分子IPP与1分子DMAPP可在法尼基焦磷酸酶(FPPS)的作用下生成法尼基焦磷酸,3分子IPP与1分子DMAPP在牻牛儿基牻牛儿基焦磷酸合酶(GGPPS)作用下形成八氢番茄红素,从而进入类胡萝卜素代谢途径(Carotenoid metabolism pathway)。本试验发现延后‘巨玫瑰’与‘瑞都红玫’VviFPPS基因均显著上调;而VviGGPPS基因均出现下调。
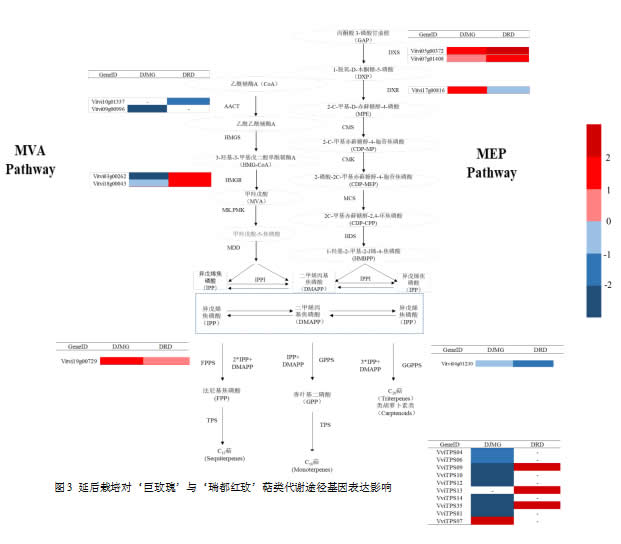
FPP、香叶基二磷酸(GPP)、GGPP在萜类合酶(TPS)作用下可形成多种的萜类骨架,FPP可在TPS催化下形成青蒿素的前体紫穗槐二烯;GPP在TPS作用下形成香叶醇、芳樟醇、橙花醇等单萜类化合物。根据序列同源性,TPS基因主要由TPS-a、TPS-b与TPS-g亚家族构成,其中TPS-b与TPS-g是产生单萜类化合物主要的修饰酶,目前TPS基因序列正得到进一步功能验证。本研究中检测到了10个VviTPS基因在‘巨玫瑰’、‘瑞都红玫’延后与正常果中发现显著差异表达,其中VviTPS04、VviTPS06、VviTPS09、VviTPS10、VviTPS12、VviTPS14、VviTPS35、VviTPS81在延后‘巨玫瑰’中显著下调,而VviTPS09、VviTPS13、VviTPS35在延后‘瑞都红玫’中出现上调,两个品种延后与正常果TPS基因表达趋势不一致。
3. 讨论
不同的生长环境胁迫下,植物体会通过改变基因表达方式来适应外界生物与非生物胁迫影响,若植物受到高温影响,其生理生化反应的动态平衡将会打破,造成植物器官功能受损或死亡。Hsp20是一种植物小分子热激蛋白,当植物体受到高温环境胁迫时Hsp20可维持植物体内蛋白稳态,由第一章可知延后果转色期前平均气温要高于正常果,而转色至成熟时期平均温度要略低于正常果的同时期,延后‘夏黑’与‘户太八号’HSP20家族蛋白基因(Vitvi13g02014,Vitvi02g01446),延后‘巨玫瑰’热激转录因子(Vitvi04g00092)均显著下调也代表在较低温度下果实受高温胁迫程度减弱。乙二醛/甲基乙二醛氧化酶(GLOX)是一类胞外氧化酶,在植物体内过氧化物酶活性过低时,GLOX可以阻止组织体内过高浓度的过氧化氢形成,延后‘户太八号’‘巨玫瑰’‘瑞都红玫’果实发现乙二醛/甲基乙二醛氧化酶基因(Vitvi02g00230,Vitvi01g00922,Vitvi18g01063,Vitvi01g00922)显著下调,可能与果实内过氧化氢浓度下降有关,有研究指出当植物体遇到盐碱或高温胁迫时,体内过氧化氢浓度出现上升趋势,由此可以推测GLOX酶基因显著下调也与延后果成熟期气温下降有关。本试验发现延后‘夏黑’GLOX酶基因(Vitvi02g00230)出现显著上调,推测可能是由于延后‘夏黑’受到了虫害或者其他胁迫影响导致。
草莓香型葡萄主要成香物质的酯类醇类化合物主要由脂肪酸代谢中的LOX代谢途径与氨基酸代谢途径合成,它们赋予了果实果香与绿叶清香风味。LOX作为脂氧合酶途径中第一阶段重要反应酶,可以亚麻酸亚油酸为底物生成氢过氧化物(9-HPO),有研究发现LOX酶活性与己醛与反-2-己烯醛含量成正相关,本试验延后果中检测到不同LOX酶基因同时存在上调或下调,而表3.2中延后‘夏黑’‘户太八号’‘巨玫瑰’己醛、反-2-己烯醛含量均低于正常果,推测可能是由于同工酶之间的反应效力存在差异,或田间修剪造成遮光条件差异导致。ADH作为一种氧化还原酶,它可通过相关催化反应实现醛类与酯类间的转换,研究认为不同ADH基因成员功能,表达模式差异较大,且在果实成熟时期ADH表达量逐渐降低这也与本试验中延后‘夏黑’‘户太八号’‘巨玫瑰’ADH基因表达差异和相关醇类物质含量结果相符。AAT基因活性与果实酯类物质含量密切相关,本试验中延后‘夏黑’、‘巨玫瑰’、‘户太八号’AAT基因均表现上调,而酯类物质含量要低于正常果(表2),可能与植物AAT底物偏好性差异有关。
为玫瑰香型葡萄带来玫瑰香特征风味的萜类物质由MVA途径与MEP途径生成。MVA途径中乙酰辅酶A先是由AACT酶缩合成乙酰乙酰基转移酶,经HMGS、HMGR催化生产甲羟戊酸,再由MK、PMK、MDD作用生成IPP与DMAPP。本试验中延后‘巨玫瑰’AACT与HMGR基因均表现下调,而延后‘瑞都红玫’HMGR基因表现上调;在MEP途径中DXS是成熟果实中控制单萜类物质含量的主要数量性状基因,DXR基因是萜烯类与类胡萝卜代谢途径上的重要酶,DXS与DXR基因在本试验延后‘巨玫瑰’与‘瑞都红玫’中表现为上调表达。综合MVA途径与MEP途径前期基因表达并不能解释延后与正常果萜烯类物质含量差异原因,需对萜类合成途径中下游基因表达进行分析。TPS基因作为分支庞大的合成酶家族,与不同类别萜类合成密切相关,其中拥有玫瑰香花香风味的香叶醇、橙花醇、芳樟醇等单萜类物质受到TPS-b与TPS-g基因调控。本试验中延后 ‘巨玫瑰’得到的绝大部分TPS基因显著下调,而延后 ‘瑞都红玫’表现为上调,符合表3.2中延后与正常果萜烯类含量差异规律,可推测VviTPS是果实单萜类物质合成途径的关键酶。
4. 小结
通过对‘夏黑无核’‘户太八号’‘巨玫瑰’‘瑞都红玫’延后与正常果进行转录组测序并对相关基因表达差异进行分析发现延后‘夏黑’有1665个基因显著下调,1264个基因显著上调;延后‘户太八号’显著下调/上调基因分别有961/1200个;延后‘巨玫瑰’显著下调/上调基因分别有1122/1530个;延后‘瑞都红玫’显著下调/上调基因分别有961/1200个。由于成熟期气温下降,四个品种延后果相较于正常果的乙二醛/甲基乙二醛氧化酶基因、HSP20家族蛋白基因、热激转录因子等有关高温胁迫相关基因显著下调。
对香气合成途径上相关反应酶基因进行表达差异分析发现草莓香型葡萄‘夏黑’‘户太八号’与‘巨玫瑰’延后果VviHPL1基因均表达显著上调,VviADH基因大部分为下调表达,VviAAT基因均表上调表达;玫瑰香型葡萄‘巨玫瑰’与‘瑞都红玫’延后果萜类合成途径相关催化酶基因表达不同:延后‘巨玫瑰’VviAACT、VviHMGR、VviGGPPS基因表达下调,而VviFPPS、VviDXS、VviDXR基因表达显著上调;延后‘瑞都红玫’VviAACT、VviGGPPS 、VviDXR基因表达下调,VviHMGR、VviFPPS、VviDXS基因上调表达。两品种VviTPS显著表达规律差异较大,延后‘巨玫瑰’得到大部分VviTPS基因呈下调表达;延后‘巨玫瑰’VviTPS09、VviTPS13、VviTPS35上调表达,其余基因并未检出表达差异。