熟期调控岗位
朱明涛 余俊 王美军 樊绍刚 白描 杨国顺
摘 要:本研究主要阐明钙处理对美人指葡萄花色苷的合成机制。美人指在花后45d ,以5 g/L CaCL2进行喷钙处理,每周喷一次,共喷三次,分别在花后45d、73d 、115d采样。研究结果发现:钙处理增加了叶片和果实中钙的含量,提高了叶片和果实钙调蛋白的活性;钙处理叶片中蔗糖转运相关基因SUC12和 SUC27的表达增强;叶片光合作用产物向果实内运输加强,提高果实内糖分含量;增加了果皮中花色素合成基因的相关表达。最终推测外源钙处理促进美人指果皮花色苷含量提高的原因有两个:(1)钙能够促进光合产物向果实内运输,为花色苷的合成提供基础物质;(2)钙能够促进钙调素活性的增加,钙调素促进花色苷合成相关的调节基因和结构基因上调表达。
关键词:花色苷;钙;钙调蛋白;可溶性糖;基因表达
花色苷是重要的多酚类物质,能够增强植物的抗逆性、抗病性,提升果蔬采后品质及货架期,并且是一种对人类健康十分有益的抗氧化类物质。葡萄果皮主要分为三种颜色,其中黑色和红色品种是由于花色苷在果皮中的积累造成的。花色苷的生物合成受生长发育和环境信号的影响,除受结构基因和调节基因的控制外,还受温度、光照、糖、激素等因素的影响。葡萄花青苷合成途径中的结构基因主要有苯丙氨酸解氨酶(PAL)、4-香豆酸辅酶A连接酶(4CL)、查尔酮合酶(CHS),查尔酮异构酶(CHI),黄烷酮3-羟化酶(F3H)、二羟基黄酮醇还原酶(DFR)、无色花色苷双加氧酶(LDOX)、O -甲基转移酶(OMT)和类黄酮糖基转移酶(UFGT)。研究报道,R2R3-MYB型转录因子影响葡萄花色苷的生物合成;在红色葡萄品种中,MYBA1转录因子可以调节UFGT基因表达,促进花色苷的生物合成。近期研究发现,钙对于植物花色苷的合成具有重要调控作用,一方面,钙能促进植物体内糖分的积累,为花色苷合成提供基质,另一方面是钙能诱导钙调蛋白(CaM)含量的增加,并与钙调蛋白结合形成Ca2+/CaM信号体系,启动第二信使系统,增加植物花色苷合成代谢途径中关键酶的活性。Shin等揭示内源性钙的变化水平可以调控蔗糖的形成,诱导花色苷的合成。此外,Xu等报道钙处理可上调花色苷的结构基因,增加总酚和花色苷含量。Peng等证实FvUGT1是草莓花色苷糖基化转移的关键酶,Ca2+/CaM通过与FvUGT1结合增加了花色苷的积累。
美人指葡萄,外观漂亮,先端紫红色,深受消费者喜爱,在葡萄果实发育和成熟阶段,常遇高温多雨天气,光照及昼夜温差都较北方不足,所以导致葡萄果实着色不良,从而降低了葡萄果实品质,因此,以美人指葡萄为试验材料具有重要意义。本课题前期研究表明在美人指葡萄转色期喷施5g/L的外源钙,可以显著提高葡萄品种美人指的花色苷含量。在此研究的基础上,研究外源施钙促进美人指葡萄果皮花色苷的合成机制,对于了解花色苷合成通路,提高葡萄栽培技术具有重要意义。
1 材料与方法
1.1 试验材料
本试验在湖南农业大学干杉葡萄试验基地进行,以12年生美人指葡萄为试验材料,采用“V”型整形,避雨栽培,常规管理 。盛花后45d,于晴天用5g/L的CaCl2+0.03%的土温8 0 喷施叶果, 以蒸馏水+0.03%土温80处理为对照,每个处理6株树,3次重复,每周喷施一次,共计三次。在花后45天(转色前1周)、73天(转色后2周)、115天(成熟期)随机取6片叶子和6串果实,每串选取15粒。取样后立即转移至液氮中冻存,带回实验室后,储存于-80℃冰箱中待用。
1.2 测定方法
叶片和果皮中总钙含量的测定参照Ortiz的方法并进行适当调整。钙调蛋白活性的测定参考Wang等人(2005)的方法,单位以Umg-1protein表示。可溶性糖的含量测定根据Sanchez等人(1998)的方法并进行适当调整。用CR-200色度计计算红葡萄色泽指数(CIRG),CIRG=[(180-h°)/(L*+C*)]。采用红色葡萄果实颜色指数的方法来评价果实的外观色泽:CIRG<2为黄绿;2<CIRG<4为粉红;4<CIRG<5为红色;5<CIRG<6为深红;CIRG>6为蓝黑色。花色苷含量的测定参考郝建军等的方法,单位以mg/g表示。用RNAprep试剂盒提取葡萄果皮中的RNA,以RNA作为逆转录模版,在带有gDNA过滤器的PrimeScript™RT试剂盒中提取,具体的引物信息见表1。
应用Microsoft Excel 2007 进行数据处理,应用SPSS 19.0 软件采用单因素方差分析和差异性分析。
2 结果
2.1 外源钙处理对CIRG和花色苷积累的影响
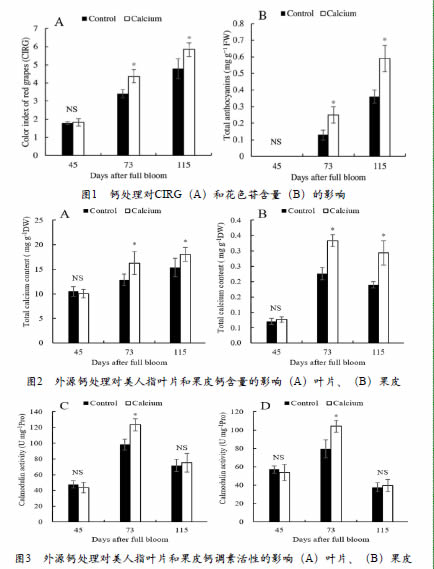
外源钙处理对美人指葡萄CIRG及花色苷含量的影响见图1。花后45天,果皮为黄绿色,花色苷暂未合成。花后73天,钙处理组CIRG为4.37,果皮由黄绿转为红色,而对照组CIRG为3.41,果皮为桃红色;钙处理组花色苷含量显著性高于对照组。花后115天,钙处理组CIRG和花色苷含量显著高于对照,钙处理组CIRG为5.84,果皮为深红色,对照组CIRG为4.79,果皮为红色。

2.2 外源钙处理对钙含量及钙调蛋白活性的影响
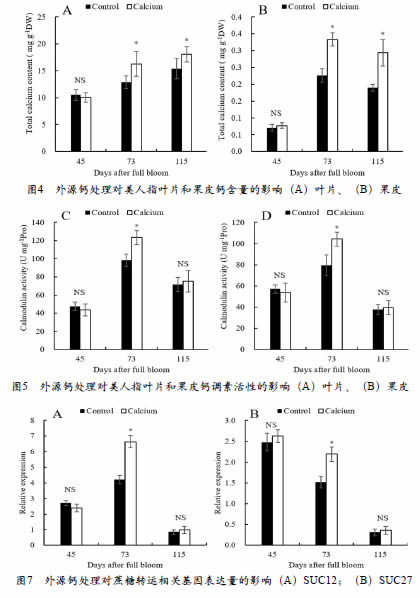
外源钙处理对美人指葡萄钙含量及钙调蛋白活性的影响见图2。花后73天、花后115天,钙处理组中叶片和果皮的钙含量显著高于对照组。在花后73天,随着钙含量的增加,钙调蛋白活性也增加。在花后115天,钙处理组叶片和果皮中的钙含量虽高于对照组,但钙调蛋白活性无显著性差异。
2.3 外源钙处理对可溶性糖和蔗糖转运相关基因的影响
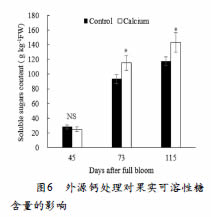
外源钙处理对美人指葡萄可溶性糖及相关表达基因的影响见图3。花后73天、花后115天,钙处理组中果实中的可溶性糖显著高于对照组。此外,还测定了与蔗糖转运相关的基因VvSUC12和V v S U C 2 7 。在花后7 3 天,VvSUC12基因表达上调,且钙处理组显著高于对照组,而VvSUC27表达水平下调,但钙处理组显著高于对照组。在花后115天时,两个基因的表达都很低,且无显著性差异。
2.4 外源钙处理对花色苷合成相关基因的影响
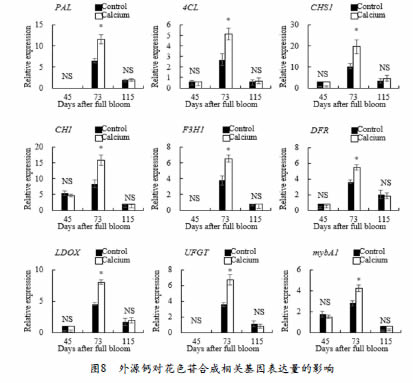
为研究外源钙处理葡萄果实中花色苷含量升高是否受到转录水平上的调控,我们对花色苷生物合成相关的结构基因和调控基因进行了检测,具体见图5。在花后73天,与花色苷合成的相关基因表达水平钙处理组显著高于对照组。在花后115天,钙处理组的所有结构基因的表达水平均下调,且与对照组无显著性差异。mybA1调节基因的表达模式相同。研究结果表明,在花后73天,外源钙处理上调了与花色苷生物合成相关的结构基因的表达水平。
3 讨论
为阐明外源钙处理对花色苷积累的调控机制,对葡萄进行喷钙处理的影响进行了相关研究。据Michailidis报道,喷钙处理可显著增加叶片和果实中的钙含量,Wang等报道钙的丰度可增加钙调蛋白的活性。我们的结果表明,在花后73天,外源钙处理显著提高了葡萄的钙含量和钙调蛋白活性。然而,在花后115天,虽然钙处理组叶片和果实中的钙含量显著高于对照组,但钙调蛋白活性没有差异。在花后73天,已经喷施了三次钙处理,高浓度的钙含量导致钙调蛋白活性增强。然而钙含量过多对细胞有害,过多的钙离子将转运到细胞壁和液泡中。因此,在花后115天,虽然叶片和果皮中的钙含量高于对照组,但大部分的钙与钙调蛋白相结合,导致钙处理组与对照组钙调蛋白活性无显著性差异。
蔗糖是高等植物中光合作用的主要产物,是碳水化合物贮藏和累积的主要形式。在葡萄果实生长发育过程中,叶片中的碳水化合物不断地转运到果实中。Pastenes研究报道,葡萄中蔗糖转运基因SUC12和SUC27的表达水平与葡萄叶片和果实中糖分的积累密切相关。
Afoufa-Bastien研究表明,蔗糖转运基因的表达受发育和环境的调控的影响。前期研究证实,钙调蛋白活性增加可调控蔗糖转运基因的表达。Shin报道,随着钙含量的增加,钙调蛋白活性升高,拟南芥蔗糖转运蛋白基因(SUC1)表达上升,增强了对蔗糖的吸收能力。
我们的研究表明,钙处理使葡萄果实叶片中钙含量增加,钙调蛋白活性增强,而且蔗糖转运基因SUC12和SUC27表达显著增强,促进叶片中的糖分向果实中转运。研究结果有助于解释钙处理为何提高了葡萄果实中的糖含量。Hiratsuka研究表明,糖是花色苷的重要成分,糖可有效促进花色苷的生物合成。Ohto报道,在拟南芥中,幼苗在含糖的环境下生长,子叶中的花色苷含量会显著积累。Michail等(2017)通过代谢组分析发现采前对甜樱桃喷施钙可以提高果实果糖、鼠李糖等的含量,而这些糖正是形成花色苷糖苷键的主要物质。在葡萄细胞和萝卜下胚轴中也有类似的报道。我们的研究结果表明,钙处理组果实中的可溶性糖和花色苷含量显著增加。
另一种可能的解释是钙处理后花色苷含量增加促使花色苷生物合成的相关结构基因表达。Gollop报道钙处理能使葡萄细胞中花色苷生物合成的DFR基因表达上调。Xu 等(2014)发现草莓成熟前喷施钙肥能够促进花色苷合成相关基因DFR、ANS和UGT的表达,并使果实花色苷含量显著增加。此外,他们还报告外源钙对花色苷结构基因表达水平的上调的作用机理较复杂。我们的研究,分析了外源钙处理对葡萄果皮中花色苷结构基因和调控基因(PAL,4CL,CHS,CHI,F3H,DFR,LDOX,UFGT和mybA1)的表达影响,结果表明在转色期,这些基因的表达水平钙处理组显著高于对照组。然则Shin等人报道,内源性钙水平的改变可以调节糖分摄取,糖信号上调促进花色素苷生物合成的结构基因和调控基因的表达。
MYB转录因子基因的表达与钙调素活性密切相关。胞内钙浓度增加,就会激活Ca/CaM系统。Yoo等(2005)在拟南芥中发现Ca/CaM可以与MYB结合,调节结构基因UFGT的表达。彭等人研究在草莓花色苷生物合成中,发现Ca与CaM结合正向调控FvUGT1活性。我们的研究结果证实了外源钙处理能增加葡萄果实中花色苷的积累,可调节花色素苷生物合成相关的结构基因和调控基因。然而,外源钙的作用可能是复杂的。花色苷生物合成的基因表达到底是糖信号或者是由钙调素诱导,目前仍不清楚,后续转录组的分析可能有助于阐明钙对花色苷生物合成的通路。
4 结论
研究数据表明钙处理增加了叶片和果实中钙的含量,提高了叶片和果实钙调蛋白的活性;钙处理叶片中蔗糖转运相关基因SUC12和 SUC27的表达增强;加强叶片光合作用产物向果实内运输,提高果实内糖分含量;增加了果皮中花色素合成基因的相关表达。最终推测外源钙处理促进美人指果皮花色苷含量提高的原因有两个:(1)钙能够促进光合产物向果实内运输,为花色苷的合成提供基础物质;(2)钙能够促进钙调素活性的增加,钙调素促进花色苷合成相关的调节基因和结构基因上调表达。