生态与土壤管理岗位
徐玉涵 翟衡 杜远鹏
摘 要: 以萌芽时期差异大的葡萄品种早巨峰和赤霞珠为材料,借助氧电极研究葡萄冬芽和根系自然休眠解除过程中总呼吸强度的变化规律及其与空气和土壤有效积温的关系,同时探求出能更准确反映葡萄休眠解除期影响根和芽呼吸变化的土壤基础温度。研究结果表明,7°C作为土壤有效积温计算的活动温度与冬芽和根系的总呼吸强度的相关性较高。与‘早巨峰’相比,‘赤霞珠’的冬芽和根系呼吸启动对有效积温的依赖性存在显著性差异(P<0.05),赤霞珠冬芽呼吸启动主要受气温有效积温的调控,根系呼吸启动主要受地温有效积温的调控。‘赤霞珠’根系呼吸启动需要的土壤有效积温高于冬芽呼吸启动需要的空气有效积温,因此根系吸收水分供应延迟,加之春季空气干燥引起的枝条水分散失可能是导致赤霞珠枝条抽干发生的主要原因之一。
关键词:葡萄,根系,芽,呼吸,空气有效积温,土壤有效积温
以早萌芽品种早巨峰和晚萌芽品种赤霞珠为试材,采用氧电极进行根系及芽呼吸进程与土壤及空气有效积温的需求关系研究,阐明土壤及空气有效积温如何影响葡萄自然休眠解除期的总呼吸进程,为探究赤霞珠抽干机理提供理论依据。
1 材料与方法
1.1 试验材料及处理
试验于山东农业大学园艺科学与工程学院试验站葡萄园基地进行。选择萌芽期晚的赤霞珠和萌芽期早的早巨峰(巨峰早熟芽变)2个品种作为研究材料。萌芽早的品种‘早巨峰’,8年生自根苗,2018在4月10号到达萌芽高峰期(萌芽率50%),萌芽晚的品种‘赤霞珠’,8年生自根苗,2018年4月25号冬芽到达萌发高峰期。两个品种南北向、相邻、平行种植,株距×行距都为2 m×3 m,单干双臂方式整形,篱架栽培。从秋季修剪以后(2017年11月22日),将温湿度探头固定在主干附近地表以下15cm处,监测期至五月初。于 2018 年2月2日至 2018 年4月25日,每10 d采一次样,每次重复 3次,每重复采集3株树共16个枝条和15cm深土层2-3mm粗的根系共20条(每条约6cm长)。每个枝条从第 3 节至第 7 节进行混合取芽,测定芽的总呼吸强度。根系剪成0.5cm根段混合取样测定总呼吸强度。本研究总呼吸强度测定时期从深休眠至萌芽高峰期。
1.2 测定指标与方法
采用路格科技温度黑匣子(L92-1)记录土壤温度。数据记录间隔为1 min。通过小型气象站(北京联创思源测控技术股份有限公司)对本园区的气温进行实时测定。按分钟计算空气有效积温和土壤有效积温(Gu., 2015),每5分钟记录一次。以10°C作为葡萄生长的气温基础温度(Molitor et al.,2014),将高于10°C的温度部分求和作为有效积温,≤10°C的温度当作0处理。计算空气日平均温度高于10℃的空气有效积温。以地表下15cm土壤温度高于7°C的温度部分求和作为有效积温,≤7°C的温度当作0处理。计算土壤日平均温度高于7℃的土壤有效积温。
利用 Oxy-Lab 氧电极通过测定溶液中氧气浓度变化来计算芽和根的总呼吸强度,略有改动(李瑾等,2009;谭钺等,2012)。改动如下:将芽和根置于密闭小瓶中,使用真空抽气机抽气,将芽及根内空气充分抽净,然后将芽及根包裹于纱布中挂在反应杯中间位置,避免芽及根与转子接触影响测定。
1.3 统计分析
采用Excel对数据进行作图并建立回归拟合曲线,采用SPSS 17.0进行相关性分析,比较总呼吸强度,显著水平P<0.05。
2 结果与分析
2.1 空气有效积温与土壤有效积温的关系分析
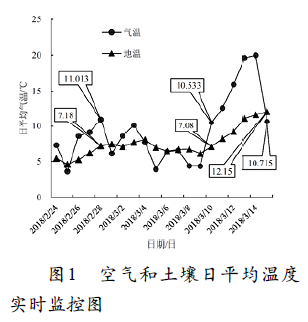
土壤有效积温与空气有效积温之间存在线性关系(孙鲁龙等,2016)。土壤温度受到空气温度的调控。以往研究表明,10°C是葡萄生长的基础温度(Molitor et al.,2014)。从图1可以看出,当空气日平均温度出现10℃时,此时的土壤日平均温度为7℃(图1)。
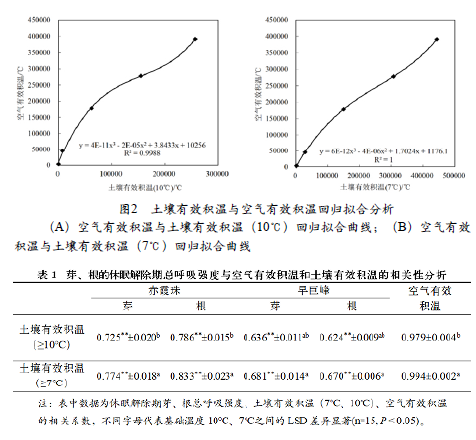
为了进一步确定以7℃作为活动温度计算土壤有效积温的有效性,以空气有效积温和土壤有效积温进行回归拟合,结果如下(图2),空气有效积温与以7℃为活动积温计算的土壤有效积温的拟合性最高(R2=1)。
为探究不同品种休眠解除期对不同土壤活动温度的活动积温的敏感性,对‘赤霞珠’和‘早巨峰’的休眠解除期总呼吸强度与不同活动温度的土壤有效积温进行相关性分析。结果如表1所示,两个品种的冬芽和根系的总呼吸强度与以7℃作为活动温度计算的土壤有效积温的相关性较高,且差异性显著。因此,本文后面的土壤有效积温都采用7℃作为土壤有效积温计算的活动温度。
表中数据为休眠解除期芽、根总呼吸强度、土壤有效积温(7℃、10℃)、空气有效积温的相关系数,不同字母代表基础温度10℃、7℃之间的LSD差异显著(n=15, P < 0.05)。
2.2 ‘赤霞珠’芽和根的总呼吸与有效积温的关系
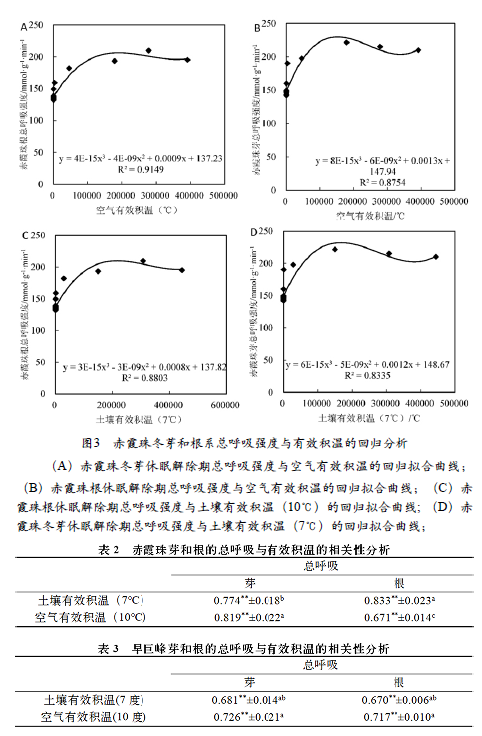
为检验有效积温对‘赤霞珠’休眠解除进度的影响, 对截至冬芽萌芽50%当天的土壤有效积温和空气有效积温进行计算, 并分别用土壤有效积温和空气有效积温与根和芽的总呼吸强度进行回归曲线拟合, 也获得了较好的拟合效果(图3-A~D)。土壤有效积温与‘赤霞珠’芽和根的拟合优度(R2)分别为0.8459和0.8819,空气有效积温与与‘赤霞珠’芽和根的拟合优度(R2)分别为0.8885和0.916, 其中空气有效积温与‘赤霞珠’芽和根的拟合度较优。
对有效积温与‘赤霞珠’芽和根的总呼吸强度之间的相关性进行分析(表2)表明,在休眠解除期间,有效积温与总呼吸强度之间都存在较好的相关性,且根和芽与有效积温的关系存在显著的不协调性。冬芽总呼吸强度与空气有效积温、土壤有效积温存在显著的相关性(r=0.819,r=0.774), 根系总呼吸强度与空气有效积温、土壤有效积温存在显著的相关性(r=0.671,r=0.833),冬芽和根系总呼吸强度与空气有效积温和土壤有效积温之间的相关性存在显著性差异。因此,‘赤霞珠’休眠解除期冬芽和根的总呼吸强度对土壤有效积温和空气有效积温都有不同的依赖性,‘赤霞珠’的冬芽的总呼吸强度对空气有效积温的依赖性较大,根系的总呼吸强度对土壤有效积温的依赖性较大。
2.3 ‘早巨峰’芽和根的总呼吸与有效积温的关系
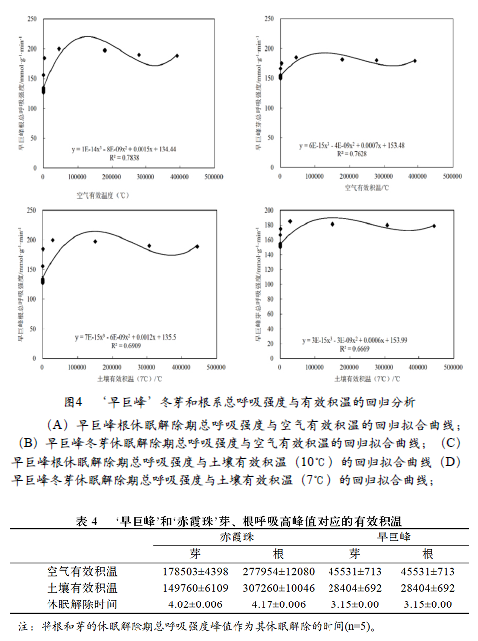
为检验有效积温对‘早巨峰’休眠解除进度的影响, 为检验有效积温对‘早巨峰’休眠解除进度的影响, 对截至冬芽萌芽50%当天的土壤有效积温和空气有效积温进行计算,将土壤有效积温和空气有效积温与‘早巨峰’根和芽的总呼吸强度进行回归曲线拟合, 也获得了较好的拟合效果(图4-A~D)。土壤有效积温与‘早巨峰’芽和根的拟合优度(R2)分别为0.696和0.697,空气有效积温与与‘早巨峰’芽和根的拟合优度(R2)分别为0.796和0.79, 其中空气有效积温与‘早巨峰’芽和根的拟合度较优。
对有效积温与‘早巨峰’芽和根的总呼吸强度之间的相关性进行分析(表3)表明,在休眠解除期间,有效积温与总呼吸强度之间都存在较好的相关性。冬芽总呼吸强度与空气有效积温、土壤有效积温存在显著的相关性(r=0.681,r=0.726),根系总呼吸强度与空气有效积温、土壤有效积温存在显著的相关性(r=0.670,r=0.717),冬芽和根系总呼吸强度与空气有效积温和土壤有效积温之间的相关性存在差异,但不显著。因此,‘早巨峰’休眠解除期冬芽和根的总呼吸强度对土壤有效积温和空气有效积温都有不同的依赖性,但其依赖性均小于‘赤霞珠’。
2.4 ‘早巨峰’和‘赤霞珠’芽、根呼吸启动时对有效积温的需求量
对‘赤霞珠’和‘早巨峰’休眠解除期的冬芽和根系的总呼吸强度与有效积温进行统计,结果如表4所示。 ‘赤霞珠’冬芽和根系对空气有效积温和土壤有效积温的需求量不同。其中,‘赤霞珠’的冬芽所需要的空气有效积温和土壤有效积温远远小于根系所需的有效积温,且其冬芽休眠解除时间比根系休眠解除提前15天。‘早巨峰’的冬芽和根系休眠解除需要的空气有效积温和土壤有效积温相同,使得休眠解除时间相一致。
3 结论
赤霞珠冬芽呼吸启动主要受气温有效积温的调控,根系呼吸启动主要受地温有效积温的调控。‘赤霞珠’根系呼吸启动需要的土壤有效积温高于冬芽呼吸启动需要的空气有效积温,早春地下部根系呼吸启动时间晚于地上部冬芽,造成早春地下部供水延迟。