酿酒葡萄品种改良岗位
唐晓萍团队
摘 要:酰基辅酶A结合蛋白(ACBP)家族基因在植物生长发育、响应逆境胁迫及生物膜修复等方面具有重要作用。本研究基于葡萄 (Vitis vinifera )基因组序列信息,鉴定并获得6个葡萄ACBP家族基因成员的全序列信息。聚类分析表明这 6个基因分属于I-IV类。依据与拟南芥的同源性比对分析,将葡萄ACBP家族基因命名为VvACBP1-VvACBP6 。通过共线性分析,发现两个VvACBP基因与拟南芥的共线性区域相对应,表明这些基因在谱系分化之前就已经存在。基因结构注释和多序列比对表明了一系列可能导致葡萄中ACBP基因功能分化的基因变异。这些信息和数据为利用 VvACBP基因家族进行葡萄抗逆性研究及应用提供了参考。
关键词:葡萄;VvACBPs;进化;非生物胁迫;
植物在生长发育过程中需要稳定的能量供应与合适的生长环境,脂类物质在该过程中发挥重要作用。脂类物质不仅为植物生长提供能量,同时也是植物体内生理和病理反应的重要参与者。ACBP家族蛋白是一类酰基载体蛋白,在脂类合成和转运过程中具有重要功能。ACBP家族具有一个高度保守的ACBP结构域,部分成员还具有锚蛋白重复序列(ankyrin repeats)、kelch- motif、coiled-coil、信号肽、跨膜域等结构域,根据以上结构域的有无将所有成员分为4类(I-IV)。Class I是一类小型ACBPs,平均由100个氨基酸残基组成,并且几乎存在于所研究的每类物种中;ClassII, 具有锚蛋白的ACBP;Class III,大型ACBPs;Class IV, 含kelch 基序的ACBP。ACBP蛋白通过调节饱和脂肪酸代谢、酰基辅酶A库、脂肪酸β氧化、囊泡运输等多种细胞内复杂的生理过程参与植物生长发育、生物非生物胁迫响应、膜修复等生理过程。
葡萄是全世界栽培面积最大、产量最高的果树,在世界及我国的果树生产中占有很重要的位置。随着气候变化及耕地面积的减少,以节省耕地、节省水资源,充分利用“阳光资源”、“山地风光”条件及山区生态多样性,推动葡萄“上山下滩”成为葡萄种植的一大选择,而高盐、干旱胁迫对葡萄生长及最终产量具有重要影响。以往的研究表明,ACBP基因家族在植物生长发育和响应胁迫方面具有重要作用。提高葡萄抗逆性、培育抗逆新品种是确保葡萄产业安全的有效途径。ACBP基因家族作为一类重要的调节植物生长发育及响应胁迫的基因家族,对其在葡萄生长及抗逆过程中的功能研究相对较少。本研究结合最近公布葡萄基因组序列信息分析葡萄ACBP基因家族结构、分布、表达特征。以期为葡萄品种选育及抗性研究提供参考依据。
1 结果与分析
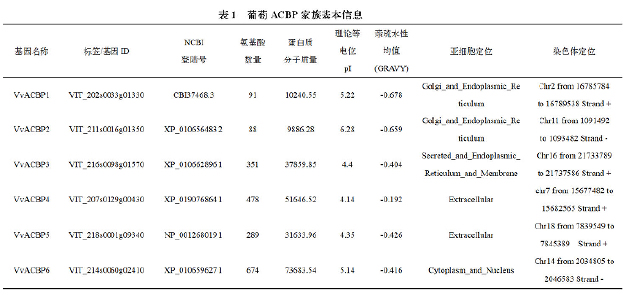
1.1 葡萄ACBP基因家族成员的鉴定及序列分析
从Pfam数据库中获得ACBP结构域的核心序列(PF00887),用HMMER筛选葡萄蛋白序列中含有该结构域的序列,共从基因组中鉴定出6个葡萄ACBP基因家族成员。对葡萄ACBP基因家族成员进行排序命名,6个VvACBP基因分别为(VIT_202s0033g01330 、V I T _ 2 1 1 s 0 0 1 6 g 0 1 3 5 0 、V I T _ 2 1 6 s 0 0 9 8 g 0 1 5 7 0 、V I T _ 2 0 7 s 0 1 2 9 g 0 0 4 3 0 、V I T _ 2 1 8 s 0 0 0 1 g 0 9 3 4 0 和VIT_214s0060g02410 )被定位到特定染色体(2、11、16、7、18 和14)。
通过ExPASy在线工具,对葡萄ACBP基因家族进行理化性质分析。分析结果显示,6个VvACBP 基因中VvACBP1 和VvACBP2 的氨基酸长度分别为 88 和 91;而VvACBP3至VvACBP6的长度范围为 289-674个氨基酸,相应分子量(MW)为9.86 kDa-73.68 kDa。其等电位范围从4.14到6.28之间,亲疏水性均值(GRAVY)范围为-0.192至-0.678(见表1)。
2.2 葡萄ACBP基因家族基因结构及系统发育树分析
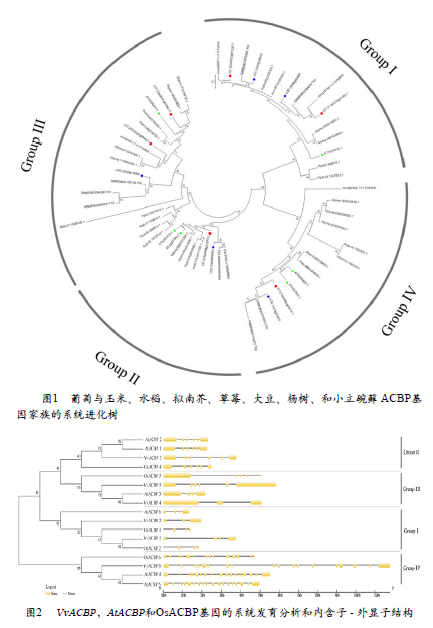
为进一步探究A C B P 基因家族的进化关系,利用ClustalXV.2.0将葡萄(VvACBP ),玉米(ZmACBP ),水稻(OsACBP ),拟南芥( A t A C B P ) , 草莓(FvACBP ),大豆(GmACBP ),杨树( A C ) 和小立碗藓(PpACBP )ACBP基因家族蛋白进行多序列比对,使用MEGA 6.0软件,采用ML(Maximum likelihood)法(Bootstrap=1 000)进行系统进化树的构建(图1)。58 个 ACBP 基因中,4 个来自草莓,5 个来自水稻,6 个来自葡萄和拟南芥,8 个来自杨树,9 个来自玉米和小立碗藓,11 个来自大豆。根据进化树的分支,可将ACBP基因分为4类,Class Ⅰ、Ⅱ、Ⅲ和Ⅳ,VvACBP 基因分布于所有分类中:VvACBP1 和VvACBP2 属于 Class I;VvACBP3具有锚蛋白重复序列,属于ClassII;VvACBP4 和 VvACBP5属于ClassIII;含 kelch 基序的 VvACBP6 属于Class IV。
为进一步研究VvACBP 基因结构的特征,用在线软件 GSDS 2.0分析葡萄、拟南芥和水稻的基因结构(图2)。结构分析表明,VvACBP 基因家族成员的基因结构复杂多变,不同的VvACBP基因所含外显子和内含子数量具有较大差异,外显子数在3-18之间,内含子数在2-17之间。其中ClassⅠ、Ⅱ、Ⅲ中的 5 个VvACBP基因只有 3-6 个外显子。Class Ⅳ(VvACBP6 )有 18个外显子(图 2)。在单子叶水稻和双子叶拟南芥中也观察到了类似的ACBP 家族成员外显子-内含子丰富/稀少模式。与Class II基因编码区内 6 个外显子相比,Class I基因编码区含有 4 个外显子,Class IV含有14 个或更多外显子,而ClassIII中没有确定数值,如 AtACBP3 含有 4 个外显子,OsACBP5 含有 5 个外显子,VvACBP4、VvACBP5 分别含有 3 个和 6 个外显子(图 2)。
比较不同组内 VvACBPs 的内含子分布,进一步揭示了 VvACBPs 的扩增方式与基因复制的产物相关,这些产物是基于早期多个内含子同源物的内含子丢失。因此,ACBP家族的扩增依赖于古代多个内含子ACBP谱系以及基因复制。
2.3 葡萄ACBP家族基因与拟南芥ACBP家族基因共线性分析
利用基因组比较研究方法,将已有的经过系统研究的物种的基因组结构和功能知识转移到研究较少的物种通常是一种方便且高效的方法。因此,我们的研究将葡萄中ACBP家族基因与模式植物拟南芥中的同源基因进行比较,以预测它们的功能。
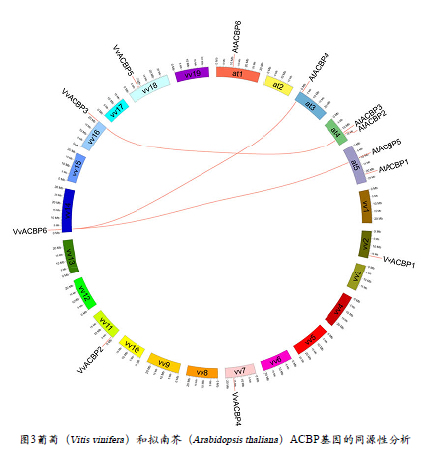
根据比较基因组分析,确定了包含两个 VvACBP和三个 AtACBP具有直系同源性(图 3)。根据该分析,三个配对的 VvACBP-AtACBP基因(VvACBP 3 -AT4G2 7 7 8 0 、VvACBP6-AT3G05420 和 VvACBP6-AT5G27630 )位于葡萄和拟南芥基因组之间具有同线性的基因组区域。这些基因可能来源于一个共同的祖先,具有潜在的相似功能。在拟南芥保卫细胞中检测到AtACBP2:GUS 的表达,并可被干旱胁迫反应和氧化应激 (H2O2) 诱导。它们在转基因植物中的过表达引起H2O2 处理后脂质氢过氧化物含量降低,并在氧化胁迫后促进膜的恢复。此外,带有 kelch 基序的AtACBP4 和 AtACBP5 通过这些结构域与伴侣蛋白相互作用并介导蛋白-蛋白相互作用。AtACBP4 还通过在角质层形成中发挥作用而有助于耐旱性。因此,葡萄的同源基因可能具有相似的功能,然而,需要进一步的实验分析来证实这一点。
3 结论
通过整个基因家族的系统发育树和功能分析是研究基因进化和功能差异的有效方法。本研究通过全葡萄基因组序列分析,挖掘到了 6个 ACBP 基因。系统分析了它们的基因组位置、基因家族特征、基因结构、基因同线性。序列分析结果表明,6 个葡萄 ACBPs 分为 4 个系统发育组。基因结构和基序分析表明,每个组都有相似或保守的内含子-外显子和蛋白质基序分布。由于该基因家族在植物中发挥的重要作用,我们期望通过进一步的研究,了解葡萄响应环境限制的分子机制,探究 ACBP在葡萄发育和形态进化中的作用。