华东华南栽培区岗位
侯旭东
陶建敏
摘要:葡萄果实采摘后,树体主要靠叶片的光合作用来合成有机物,贮存营养。果实采收后结果枝短截,增加树体通风透光,促进基部叶片光合作用。本研究以‘夏黑’葡萄为研究材料,在葡萄采摘后,对一年生结果枝进行不同程度的短截,测定分析不同短截程度对葡萄结果枝基部叶净光合速率、气孔导度、胞间CO2浓度及蒸腾速率的影响。结果表明:与对照(CK)相比,不同的短截程度都明显提高了‘夏黑’葡萄基部叶片的净光合速率(Pn)、气孔导度(Gs)及蒸腾速率(Tr),有利于基部叶片的光合积累,而胞间CO2浓度(Ci)有所降低。结论:短截对‘夏黑’葡萄基部叶片光合有明显的促进作用,留4片叶短截促进作用最为明显。
关键词:结果枝;短截;‘夏黑’葡萄;光合特性
‘夏黑’葡萄(Vi t is v i n i f er aL.×Vi tis labrusca L.)原产于日本,欧美杂交种,三倍体品种。在1968年,由日本山梨县果树试验场利用巨峰和二倍体无核白杂交育成,1998年引入我国。‘夏黑’葡萄果粒近圆形,自然粒重3.5克左右,早熟,无核,高糖低酸,香味浓郁,硬度中等,可溶性固形物在20%左右。‘夏黑’葡萄因有丰产、抗病和耐贮运等优点,深受消费者喜爱,种植面积在逐年扩大。并且人们生活水平的提高,促使对葡萄品质的要求也越来越高,优质高效的栽培技术就显得尤为重要。
短截是指对一年生枝条进行剪短,留下部分枝条进行生长。在生产上,短截常用于树冠中骨干枝延长枝修剪,特别是幼树的整形修剪,能明显的增加分枝量,减少枝条的“光腿现象”。短截在维护树势上下均衡、矫正树形等方面有重要作用。短截分为轻短截、中短截、重短截和极重短截这4种程度。果树的库源关系变化对果树产量和品质影响很大,果树的一些生产栽培技术措施如疏花疏果、夏季修剪、短截等均是基于此原理。果树产量主要依赖于叶片光合同化有机物,然后转运分配到果实中,有机物的转运分配主要取决于库源相互作用。果树源与库之间有着复杂的联系,研究库源关系与光合作用的影响能为更有效的调节果树的光合能力提供理论依据。
光是所有绿色植物光合作用中光能的唯一来源,植物干物质的90%以上来自叶片的光合作用。叶片光合能力的大小和叶片光合作用功能时期的时间长短是影响植物生长发育的决定性因素,同时还会影响果实品质,进而影响经济价值产量;叶片的叶绿体不仅受植物本身的生长时期的影响,而且还受很多外界因素的影响。果树的光合作用受诸多因素影响,崔春梅等和艾沙红•买买提等研究表明短截也是影响果树的光合作用的一个重要因素,所以研究不同短截程度的光合特性,对于指导葡萄生产有很大的实际意义。
1 实验材料与方法
1.1 实验材料
实验于2015年南京农业大学汤山葡萄试验基地进行,平棚架“H”型整形,避雨栽培,南北走向,行株距为6.0 m×3.0 m。试验品种‘夏黑’葡萄,选择9棵4年生长势中庸相近的植株,通风、透光条件良好,沙壤土土质,采用滴灌技术,管理水平较高。
1.2 实验设计
葡萄采摘过后,随机选取树龄、长势和叶片发育程度等相似的‘夏黑’葡萄树9棵。在9棵树上分别选取5枝(1枝备用)长势、叶面大小及叶片朝向(特别是基部第三个叶片)等生理特征相似的一年生外延枝条作为实验材料。每株树5枝外延枝条,分别作留4、6、8片叶子和CK共四个处理(剩余1枝作为备用)。然后把9棵树随机分为3组,每组3棵树,分别为a组,b组,c组。a组为实验组,b组和c组为重复。
1.3 测定方法
果实采摘后对一年生结果枝进行短截,短截处理7d后,选择晴天9:30-10:30对基部叶片进行光合特性的测定。选用美国LI-COR公司生产的LI-6400便携式光合测定仪测定。在测量光合速率之前先用干净的抹布擦去所有待测叶片表面的灰尘(减少实验误差),然后调试仪器进行测定,选取各处理结果枝基部第三片叶片(叶面大小、朝向等相似)作为测定对象,测定各处理选取叶片的净光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)、蒸腾速率(Tr),每个叶片重复3次,取平均值。流速设定为500μmol•s-1,根据当时大棚光强设定红蓝光源光强为1000μmol•m-2•s-1。每隔7d重复测定一次,共测定5次。在最后一次光合特性测定时分别取8点、10点、12点、14点和16点五个时间点进行光合日变化的测定。
1.4 数据处理
采用SPSS和Excel软件进行数据统计和绘图。
2 结果与分析
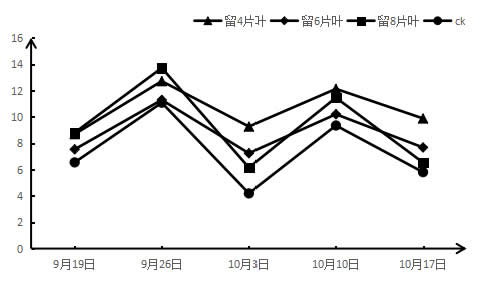
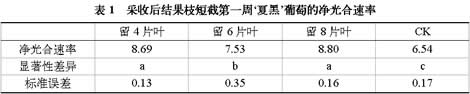
图1 采收后结果枝短截对‘夏黑’葡萄净光合速率的影响
2.1 采收后结果枝短截对‘夏黑’葡萄净光合速率的影响不同短截处理叶片净光合速率如图1所示,从总体趋势来观察留4片叶处理三个时期的净光合速率高于其他处理,并且所有处理的净光合速率都高于CK,并且所留叶片越少,净光合速率基本也就越大,基本呈现为:留4片叶>留6片叶>留8片叶>CK。
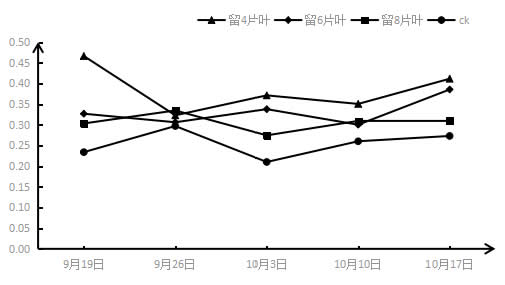
图2 采收后结果枝短截对‘夏黑’葡萄胞间二氧化碳浓度(Ci)的影响
如表1所示,短截处理一周后不同处理间的净光合率具有一定差异,短截处理的净光合速率与CK具有显著性差异,留6片叶和其他处理也具有显著性差异,而留4片叶和留8片叶之间的显著性差异不明显;第二周处理时长和温度等因素都适宜,所有处理的净光合速率都达到最大,第三周时净光合速率差异最为明显,所测老叶进行光合作用的能力也被完全激发,净光合速率也符合“所留叶片越少净光合速率越高”的总体趋势;在第四周之后,所有叶片的净光合速率都开始有所下降。
总的来说,短截处理后基部叶片的净光合速率基本呈先上升后下降的趋势,但始终比第一周的净光合速率高。第三周的净光合速率不符合“先上升后下降”的规律,留8片叶处理在第二周和第五周都不符合“所留叶片越少净光合速率越高”的总体趋势。
2.2 采收后结果枝短截对‘夏黑’葡萄气孔导度Gs的影响如图2所示,各个处理在不同时期都具有一定的差异,处理第一周时,留4片叶>留6片叶>留8片叶>CK;在第三周和第五周时,也是留4片叶>留6片叶>留8片叶>CK。随着时间的推移,基部叶片的气孔导度总体上呈现上升的趋势,与净光合速率的速率呈相反的趋势,但不同短截处理的Gs都始终高于CK。大多数枝条短截程度越大,气孔导度也越大,总体呈正相关趋势,但第二周和第四周的留8片叶处理不成正相关。
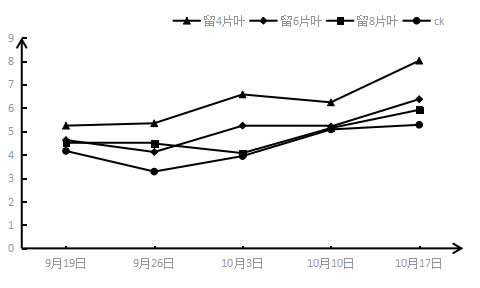
图3 采收后结果枝短截对‘夏黑’葡萄气孔导度
(Gs)
的影响
2.3 采收后结果枝短截对‘夏黑’葡萄蒸腾速率Tr的影响如图3所示,短截处理过后,所有短截处理叶片的蒸腾速率都呈上升趋势,与气孔导度的上升趋势相同,即处理时间越长基部叶片的蒸腾速率越大,但是与净光合速率的趋势有所差异。在不同短截处理之间,基本呈现所留叶片越少叶片蒸腾速率越大的状况,即留4片叶>留6片叶>留8片叶>CK,能观察到所有短截处理叶片的蒸腾速率Tr都大于CK,且留4片叶处理的Tr最大。但在第二周的留8片和6片叶子处理不满足此规律。
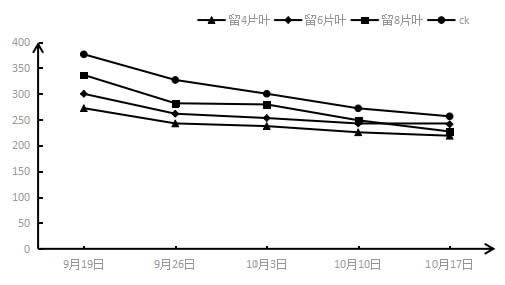
图4 采收后结果枝短截对‘夏黑’葡萄蒸腾速率(Tr)的影响
2.4 采收后结果枝短截对‘夏黑’葡萄胞间二氧化碳浓度Ci的影响
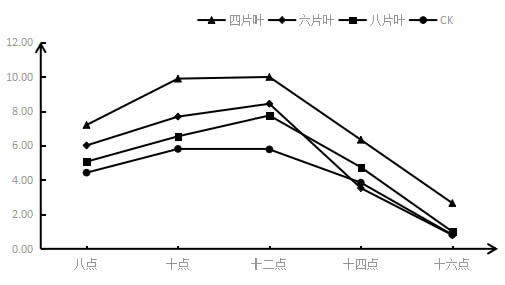
图5 采收后结果枝短截对‘夏黑’葡萄胞间二氧化碳浓度(Ci)的影响
如图4所示,所有短截处理叶片的胞间CO2浓度都随着处理时间的推移呈下降的趋势,正好与叶片气孔导度和蒸腾速率的趋势完全相反,与叶片净光合速率的趋势相似,但也有很多不同之处。叶片的胞间CO2浓度是呈直接下降趋势,而叶片的净光合速率是呈先上升再下降的趋势。在不同处理之间,短截程度越大,叶片胞间二氧化碳浓度越小,所有短截处理的胞间CO2浓度都低于CK,其中留4片叶处理的叶片胞间CO2浓度最低。但在第五周时,留8片和6片叶子的胞间CO2浓度不符合此规律,而且各短截处理叶片的胞间CO2浓度差异不明显。总体上看各处理叶片的胞间CO2浓度,CK>留8片叶>留6片叶>留4片叶。
2.5 采收后结果枝短截对‘夏黑’葡萄光合作用日变化的影响
图6 采收后结果枝短截对‘夏黑’葡萄光合作用日变化的影响
如图5所示,所有短截处理的光合作用日变化都是先上升后下降,因为这与光照强度有关。叶片的光合速率与光照强度成正比,光照先增强后减弱,即所测的光合作用速率也是先增大再减少;短截处理的光合作用日变化曲线基本都在CK的上方,留4片叶处理的光合作用日变化曲线在最上方,光合作用最强。排列顺序基本上是:留4片叶>留6片叶>留8片叶>CK。
3 讨
论
短截技术在果树上应用较为普遍,在葡萄上也比较重要。衰老树采用回缩和短截等更新修剪方法,调节树体营养与水分的分配运转,改善树体营养状况,提高叶片的光合速率。植物光合作用主要是通过叶片进行的,叶片性状的差异是造成光和差异的主要原因之一,叶龄和叶位与光合作用关系也很密切,许多在果树上的研究表明,净光合速率由于叶龄和叶位的不同而出现差异。所以我们的测量对象全部选择为生长状况相似的一年生结果枝基部第三个叶片,还尽量选择叶片朝向和位置等相似的叶片,最大程度的减少了外界环境对叶片光合作用的影响。光合作用是植物生长发育和产量品质形成的基础。叶片光合速率随着叶龄的变化而变化。果树单叶光合速率变化,一般规律是叶片初展尚呈绿黄色时,净光合速率很低,随叶片生长,净光合速率提高,叶片定形时,净光合速率最高,维持一段时间,随叶片衰老,净光合速率降低。
本实验结果表明,经短截处理的净光合速率都显著高于CK的净光合速率。其中第三周所有叶片的净光合速率都偏低,可能是测量当天的天气是多云,并且温度也偏低,因为光线和温度是影响净光合速率的重要原因。测量叶片的净光合速率随着处理时间的推移呈现先升高后下降的趋势,老叶光合作用加强后促进了自身的老化可能是导致短截过后叶片光合能力下降的主要原因之一,叶片内部分机制退化后导致净光合速率下降;天气转凉,温度降低,也会导致光合速率降低;还有可能是天气的变凉的,导致各种酶活性降低等原因。总体来说,叶片的净光合速率先升高后下降,且随着短截程度的增加净光合速率也随之升高,即:留4片叶>留6片叶>留8片叶>CK。净光合速率是光合特性的一个重要组成部分,它能直接反映叶片积累同化物的能力,而植物基部叶片的光合作用受短截处理的影响,不同的短截程度会导致基部叶片净光合速率不同,从而影响基部叶片积累有机物的能力。本研究证明通过短截处理能够明显的促进基部叶片的净光合速率,并且在一定程度下短截程度越大,促进效果越明显。
与净光合速率不同,叶片的气孔导度和蒸腾速率随着处理时间的推移都基本呈现上升的趋势。不同程度短截处理的气孔导度及蒸腾速率都明显高于没有进行短截处理(CK),并且短截程度越大,叶片的气孔导度和蒸腾速率也越大,即:留4片叶>留6片叶>留8片叶>CK。但留8片叶子处理在第二周和第四周的气孔导度不符合这个趋势,以及留8片叶子处理在第二周的蒸腾速率也与此趋势不相符,可能是由于此处理选取的叶片具有一定的特殊性(如:叶片朝向和位置等)。叶片的胞间CO2浓度却与气孔导度和蒸腾速率完全相反,叶片的胞间CO2浓度随着处理时间的推移都基本呈现下降的趋势,不同程度短截处理的胞间CO2浓度要低于没有进行短截处理的CK,呈现的趋势为:CK>留8片叶>留6片叶>留4片叶。留8片叶子处理在第五周的胞间CO2浓度与此规律不相符,原因可能与上文气孔导度和蒸腾速率相同。叶片的光合作用日变化曲线呈现为先上升后下降的“单峰曲线”,与文晓鹏等和张国良等报道的双峰有所不同,可能是在进行叶片光合作用日变化测量时气候开始转凉,日最高温度不是太高,中午不出现“午休现象”,从而只能形成单峰。
本研究证明短截有利于‘夏黑’葡萄基部叶片的光合积累,且在一定范围内短截程度越大,促进效果越明显。本实验留4片叶处理最有利于‘夏黑’葡萄基部叶片的光合积累。短截促进叶片光合速率的原因可能是短截后只有几个芽在剪口下抽出枝条,改变了库源比,减少了叶幕,增加了对存留枝梢叶片生产光合产物的需要,而且也减少与来自根系的细胞分裂素和无机营养的竞争,从而使存留叶片的光合速率提高。
据Zhou等和Iglesias等报道,改变植物光合产物的库源比之后能观察到光合速率的变化。去掉部分叶片来提高库源比会导致源中碳水化合物含量下降,不仅增加对存留叶片生产光合产物的需要,而且也减小了对来自根系的细胞分裂素和无机营养的竞争,从而使存留叶片的光合速率提高。本实验在葡萄采摘后,对一年生外延枝条进行不同程度的短截,除去外延枝条上的嫩叶,相当于减少了源的个数,从而使促进剩下的源增强工作,进而能够促进基部叶片光合作用。
Palit利用14C同位素技术研究发现, 顶部叶片会将光合产物分配给顶芽和幼叶,繁殖器官会从就近的叶片获取光合产物。郭太君等在葡萄的光合产物分配研究中得出果实膨大期形成的同化产物积累在果实中的比例高于开花期和成熟期,且同化产物具有就近分配的特点。李合生等提到叶片进行光合作用所产生的营养物质会依照“就近原则”进行转运和分配。根据同化物就近分配的特点,基部叶片光合作用的提高有利于同化物的积累,并且就近分配给基部的芽,外延枝条基部的芽会储存更多的有机物质更加饱满,进而能够促进枝条基部芽花芽形成,有利于下一年的花芽萌发,还有利于简化树形采用冬季短梢和极短梢修剪。
4 结
论
综上所述,不同程度短截处理与对照(CK)相比,均有效的提高了基部叶片的净光合速率、气孔导度及蒸腾速率,降低了胞间 CO2浓度。所以短截处理能促进叶片的光合作用,有利于‘夏黑’基部叶片的光合积累,在一定范围内短截程度越大,越有利于光合产物的积累,本研究中留4片叶处理促进效果最明显。基部叶片光合能力增强,光合作用会积累更多的同化物就近分配给基部的芽,枝条基部的芽就能储存更多的有机物质更加饱满,进而促进基部芽的花芽形成,有利于下一年花芽萌发。因此,通过短截处理技术来提高基部叶片的光合产物的积累是可行的,值得进一步研究应用,我们还可以在此技术的基础上研究短截技术对下一年花芽萌发的影响。