华东华南栽培岗位
王继源 辛守鹏 陶建敏
细胞分裂素( CPPU)是葡萄实际生产中最常用的生长调节剂之一,CPPU的生理作用是促进细胞分裂和扩大,加快叶绿素合成,增加叶片光合效率,调配光合营养产物的分配比例,延缓衰老,提高对胁迫条件下的抗逆性,因此,CPPU可以明显增大葡萄果实大小,对提高果粒质量与产量的作用明显。通常认为葡萄果实的成熟是由脱落酸调控的,而脱落酸(ABA)一直以来都被认为在果实成熟衰老过程中起到重要作用,它在高等植物中的合成器官有老叶、根尖、花、果实和种子,葡萄果实中的脱落酸是由叶片和果实自身合成。脱落酸的生物合成途径有直接途径和间接途径,间接途径是高等植物合成ABA的主要途径,9-顺环氧类胡萝卜素双加氧酶(NCED)是其中的关键酶,而βG1基因是调控ABA由结合态向游离态转变的重要基因。因此,根据实际生产与研究需要,研究了不同浓度细胞分裂素(CPPU)处理果穗对不同叶位叶片光合特性及果实ABA合成相关基因VvNCED1、VvβG1表达量影响,对解决葡萄发育成熟存在的问题有重大意义。
1 材料与方法
1.1 试验材料
试验于2014年4—10月在南京农业大学汤山葡萄试验基地进行。平棚架“H”型整形,避雨栽培,南北走向,行株距为6.0m×3.0m。试验品种 ‘阳光玫瑰’葡萄,选择长势中庸相近的植株,将东侧枝条基本一致的单花穗挂牌标记,以常规方法进行田间土肥水管理及病虫害防治。
1.2 试验方法
处理前对花穗进行修剪,除去穗肩小穗,留穗尖长约4cm,花后一周分别用清水(CK)、10 mg•L-1CPPU(处理A)、20 mg•L-1 CPPU(处理B)处理果穗5s,每处理3棵树。
处理4周后,选择晴天9:30-10:30,从标记的枝条中选取长势一致无其他叶遮挡的枝条,用美国LI-COR公司生产的LI-6400便携式光合测定仪分别测定每根枝条从基部数第4叶(下位叶,果穗邻近叶)、第8叶(中位叶)、第12叶(上位叶)片叶净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)等相关指标, 重复3 次, 取平均值。流速设定为500μmol•s-1,根据当时大棚光强设定红蓝光源光强为1000μmol•m-2•s-1。测定之后开始采集果粒样品,每隔1周取样1次。
取样时脱脂棉拭去果粒表皮异物,用手术刀迅速将果皮与果肉剥离,锡箔纸包好,分装两份,放入液氮中带回实验室,-70℃保存备用。采用改良CTAB法分别提取葡萄果皮与果肉中的总RNA,以总RNA为模板, 利用Ta k a r aPrime ScriptTM RT-PCR试剂盒反转录合成cDNA。根据Genbank中VvNCED1、VvβG1、Vvactin1的全长序列,利用Primer5.0设计PCR引物(引物序列见表1),引物由上海捷瑞公司合成。以 cDNA为模板,Vvactin1基因作为内参,利用合成的引物进行Real-Time PCR扩增检测相关基因的转录水平。PCR反应体系为应20μL:含有SYBRPremix Ex Taq(2×)10μL,上游引物和下游引物各0.2μL,模版1μL和8.6μL去离子水。反应程序如下: 95℃预变性90s,95℃变性15s,60℃退火20s,72℃延伸40s,40个循环。每个模板做 3个平行,用平均值表示总RNA的CT值,采用△△CT法对荧光定量PCR扩增数据进行处理,目的基因的相对含量通过计算 值来确定。
1.3 数据分析
试验数据采用Excel进行统计和绘图,用SPSS软件进行方差分析。
2 结果分析
2.1 不同浓度CPPU处理对葡萄叶片Pn的影响
由图1 可以看出, 不同浓度CPPU处理叶片净光合速率是中位叶>上位叶>下位叶,在CPPU处理条件下,不同叶位叶片净光合速率随着处理CPPU浓度的增加而增大,即处理B>处理A>CK,但不同叶位在CPPU处理后的净光合速率差异程度不同,差异程度为下位叶>中位叶>上位叶,且上位叶不同处理间差异不显著。
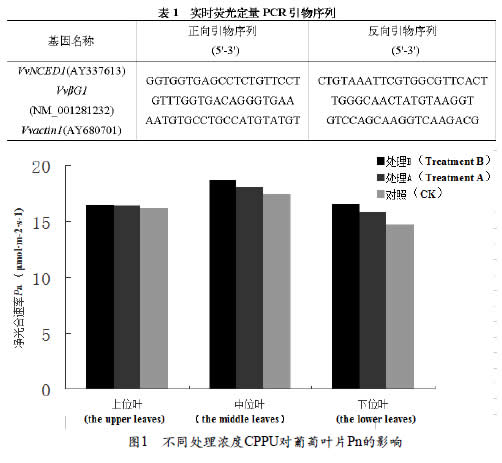
2.2 不同浓度CPPU处理对葡萄叶片Gs的影响
由图2 可以看出, 不同浓度CPPU处理气孔导度是中位叶>上位叶>下位叶,在CPPU处理条件下,不同叶位叶片气孔导度随着处理CPPU浓度的增加而增大,即处理B>处理A>CK,但不同叶位在CPPU处理后的气孔导度率差异程度不同,差异程度为下位叶>中位叶>上位叶,且上位叶不同处理间差异不显著。
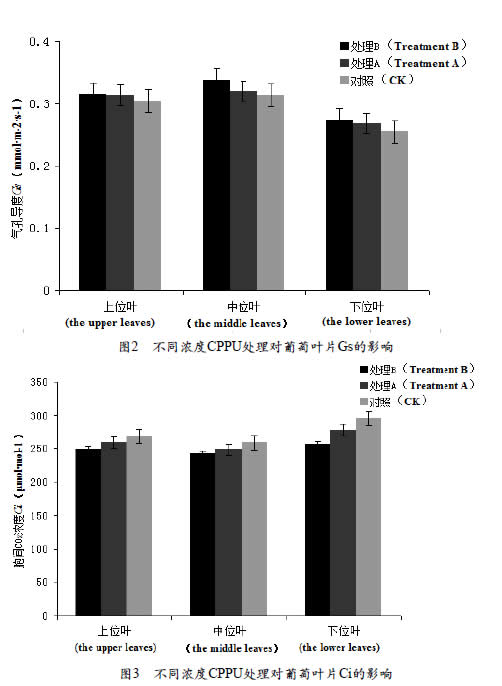
2.3 不同浓度CPPU处理对葡萄叶片Ci的影响
由图3 可以看出, 不同浓度CPPU处理胞间二氧化碳浓度是中位叶<上位叶<下位叶,在CPPU处理条件下,不同叶位叶片胞间二氧化碳浓度随着处理CPPU浓度的增加而降低,即处理B<处理A<CK,但不同叶位在CPPU处理后的气孔导度率差异程度不同,差异程度为下位叶>中位叶>上位叶,且上位叶不同处理间差异不显著。
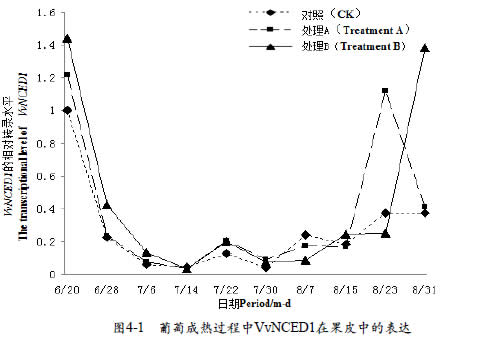
2.4 不同浓度CPPU处理对葡萄果实VvNCED1基因转录水平的影响
实时荧光定量P C R 对VvNCED1基因表达分析显示,果皮中(图4-1)VvNCED1在幼果期到转色期的过程中,基因表达量呈逐渐降低趋势,转色期期间表达量小幅增加,有个小峰,转色期过后VvNCED1基因表达量呈逐渐增加的趋势;不同处理间存在较大差异,CPPU处理有利于转色期之前VvNCED1基因表达量增加,转色期期间各个处理VvNCED1基因表达量差异不明显,转色期之后CPPU处理也有利于VvNCED1基因表达量增加,但却将大量表达时间推迟了2周左右。果肉中(图4-2),CK中基因VvNCED1在整个生长发育过程中表达量都很低,但CPPU处理提高了VvNCED1基因在幼果期和转色期的表达量,且随CPPU浓度增加增大越明显。
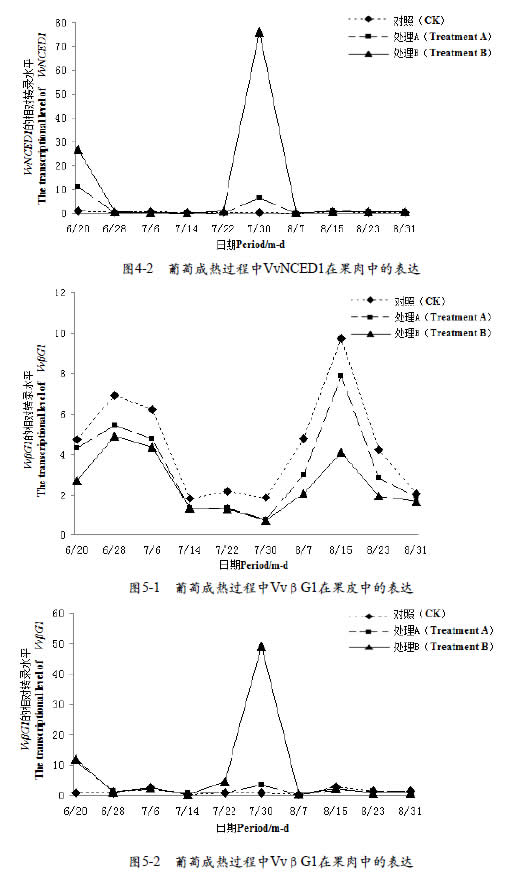
2.5 不同浓度CPPU处理对葡萄果实VvβG1基因转录水平的影响
实时荧光定量PCR对VvβG1基因表达分析显示,果皮中(图5-1),在幼果期和膨大期VvβG1基因都有较高的表达量,但CPPU处理会抑制VvβG1基因在这两个时期的表达,转色期时,各个处理VvβG1基因表达量没有明显差异,转色期之后,CK中VvβG1基因表达量稍高于CPPU处理。果肉中(图5-2)CK中基因VvβG1在整个生长发育过程中表达量都很低,但CPPU处理提高了VvβG1基因在幼果期和转色期的表达量,且随CPPU浓度增加增大越明显。
3 讨论
利用植物生长调节剂来调控果树生长发育已是生产中的重要技术措施,CPPU作为细胞分裂素类生长调节剂被广泛应用于葡萄实际的生产中。葡萄幼果期使用CPPU对葡萄果实有明显的增大作用。由于果实中的有机物绝大部分来自于叶片的光合作用,因此,葡萄果实增大则库强增大,必然对源叶片产生巨大的影响。作为库的果实和幼叶对源的光合产物都具有竞争力,本研究表明,果实库强增大使叶片光合作用增大,且随着距离果实叶位距离的增加叶片光合作用受到的影响而减小。上位叶的光合产物主要是就近分配给幼叶,且距离果实较远,所以其几乎不受果实库力变化的影响。下位叶成熟较早,而出现一定程度的衰老,所以光合效率明显低于中位叶,但受到库源关系的影响程度较大。随着CPPU处理浓度的增加,各处理叶的净光合速率、气孔导度明显增加,胞间CO2浓度降低.光合作用的增加有利于CO2的固定,从而产生更多的有机物。用CPPU处理幼果,可以明显增强相应枝条上叶片的光合作用能力。
葡萄果实成熟过程中的转录水平,发现NCED基因的表达与ABA积累几乎是同步的,CPPU处理巨峰葡萄幼果后发现CPPU先促进ABA含量升高,后又推迟ABA高峰的出现,这与本试验果皮中NCED基因表达趋势基本一致。CPPU抑制花青素的合成,提高果皮叶绿素含量,延迟果皮的退绿。本研究结果表明,CPPU处理明显抑制果皮中VvβG1基因的表达量,这说明CPPU处理会抑制ABA由结合态解离成游离态,进而延缓果皮中ABA的积累量,延缓果皮叶绿素的衰退,可能也是CPPU处理后葡萄果皮出现“滞绿”现象的原因。