南宁综合试验站
成果 周思泓 刘金标 冯启燕 韦荣福 余欢 王博 张瑛 白先进
摘 要 :亚热带地区气候条件是一年两收栽培推广和应用的基础。不同的温度、光照和水分条件是造成葡萄初生代谢物和次生代谢物差异的根本原因。我们对‘巨峰’葡萄进行了基于超高效液相色谱-三重四极质谱法(UPLC-QQQ-MS)的代谢组学研究。共鉴定出1062个代谢产物,分为10个不同类别,其中类黄酮类代谢产物最多,达285个。同一生长季,不同发育阶段葡萄的代谢产物有876种,夏季和冬季葡萄共有551种差异代谢产物。根据检测到的代谢物,分层聚类分析(HCA)和主成分分析聚类分析(PCA)能清楚区分发育和生长季节。值得注意的是,黄酮类化合物是造成浆果在生长和发育季节成分差异的最重要的化合物。大部分黄酮类化合物在冬季葡萄中含量较高,但在夏季葡萄中也有一些含量较高,如山奈酚-4-O-葡萄糖苷、无色花青素和花青素-3-O-葡萄糖苷。此外,杨梅素-3-O-阿拉伯苷在所有四个发育阶段在冬季葡萄中始终高于夏季葡萄。极高的温度和较高的相对湿度是夏季葡萄中黄酮含量低于冬季葡萄的重要原因。此外,冬葡萄发育早期较强的光照强度对黄酮类化合物的积累有积极作用,尤其是黄酮醇和黄烷-3-醇。本研究为两季葡萄中黄酮类化合物的代谢提供了新的认识。
关键词:广泛靶向的代谢组学;一年两收;类黄酮;葡萄
中国南方热带地区由于低温不足和雨季炎热,传统上被认为是不适合种植的地区。葡萄一般难以促进发芽,花芽分化缓慢,果实品质差,成熟期难以集中销售。2003年以来,一年两熟栽培技术体系的研究与应用,解决了上述传统问题,在广西形成了特色优势葡萄栽培区。葡萄一年两熟栽培技术包括利用老熟枝条冬芽的夏季促进花芽的二次分化。在第一个生长季节收获的葡萄称为夏季葡萄,在第二个生长季节收获的葡萄称为冬季葡萄。
黄酮类化合物是葡萄中重要的次生代谢产物,在葡萄生长发育、抗紫外线和抗病虫害、果实色泽、风味品质和营养价值等方面发挥着重要作用。花青素、黄酮醇和黄烷-3-醇是葡萄浆果中主要的黄酮类化合物。除了各种因素外,气候条件和栽培技术也会影响类黄酮的分布和含量,它们相互作用以调节葡萄中类黄酮的代谢。对于同一品种,气候和天气条件可以发挥非常重要的作用。光是影响黄酮类化合物代谢的最重要的气候因子,包括光照强度、光照品质和光照周期。一般来说,光照会增加类黄酮的含量,特别是花青素和黄酮醇。因此,在实践中,摘叶和套袋通常被用来控制葡萄暴露在光下的情况。温度是影响黄酮类化合物合成的另一个重要环境因素。一般来说,低温可以诱导类黄酮合成相关基因的表达,并增加其积累。然而,高温严重抑制了黄酮类化合物的合成。水分状况是影响黄酮类化合物合成的第三个重要环境因素。适当的水分缺乏增加了花青素和原花青素的积累,通过减少果实体积改善了果实,并直接上调了类黄酮合成途径相关基因的表达。因此,调节亏水灌溉可以作为增加葡萄中类黄酮含量的一种策略。
独特的气候条件是发展两季作物种植的基础。下半年光照充足,秋冬季节昼夜温差大,更有利于光合产物的积累,葡萄的花青素含量较高,色泽和品质较好。我们研究团队前期的研究表明,冬葡萄比夏葡萄具有更高的可溶性固形物含量和可滴定酸含量。我们之前利用转录组测序技术来探索两季果实类胡萝卜素和苯丙烷-类黄酮类化合物代谢途径的差异。此外,类黄酮代谢途径的某些结构基因在成熟期的冬葡萄中表达更高。先前的一项研究表明,冬季葡萄的酚类物质和香气物质比夏季葡萄更丰富。然而,基于高通量、高灵敏度和广泛覆盖的分析方法检测两种作物葡萄中的代谢物的报道很少。
采用超高效液相色谱-质谱联用技术(UPLC-QQQ-MS)分析了‘巨峰’葡萄(Vitis labrusca L.×Vitis vinifera L.)夏季和冬季葡萄中的黄酮类化合物。重点分析了气候因素对两季葡萄中黄酮类化合物含量的影响。本文对夏季和冬季葡萄中黄酮类化合物的代谢进行了研究。这为进一步研究一年两收栽培条件下葡萄风味物质的代谢机理奠定了理论基础。
2 材料和方法
2.1 葡萄园条件和一年两收的葡萄栽培方法
实验于2020年在位于南宁的广西农业科学院葡萄与葡萄酒研究所的葡萄园(北纬22 ° 51′29′ ,东经108 ° 14′33′)进行。葡萄园里的自根苗葡萄是8年生的巨峰葡萄,用单干棚架种植。葡萄行间距为2.0 m×6.0 m,采用避雨栽培模式。葡萄园的日常管理,包括病虫害和水控制方法,是按照以前的标准进行常规管理。
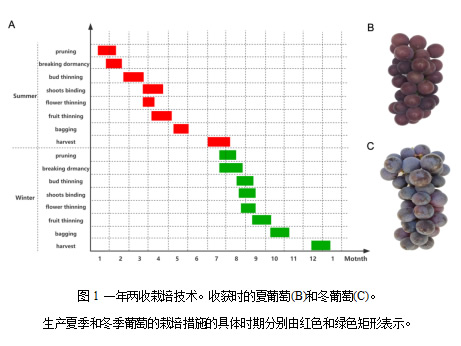
一年两收栽培的主要栽培方法如下(图1a):在一月份中长梢修剪(4-7个芽) ,然后在一月下旬至二月中旬,当温度稳定在10 °C 以上时,用50%的单氰胺稀释20次,打破休眠。花疏在三月底到四月初进行,夏季葡萄的收获(图1B)发生在六月中旬到七月初。然后,施肥是在夏季葡萄收获后进行的,以恢复葡萄藤的生长。长梢修剪(8-10个芽)发生在7月下旬至8月中旬,同时,所有的叶子被移除,休眠被打破,使用50%的单氰胺稀释30次。然后,在发芽后5-8天,一年的第二个生长周期开始,冬葡萄的收获(图1C)发生在12月下旬至1月中旬。
2.2 气象调查
2020年浆果生长过程中的气象数据,包括光照持续时间(h)、温度(°C)、相对湿度(%)和太阳辐射强度(W/m2) ,是使用Hobo temp/RH/SR智能传感器(Onset Computer Corporation,Bourne,MA,USA)收集的。每隔5分钟进行一次测量。
2.3 取样方法及理化指标监测
600粒浆果在E-L的四个阶段被采样: 浆果仍然坚硬和绿色(E-L 33) ,转色(E-L 35) ,转色结束(E-L 37) 和采收(E-L 38)。每个生物重复包含9个株,每个生物重复200粒浆果。随机选取100个浆果,测定颜色参数、浆果鲜重、可溶性固形物浓度(SSC)和可滴定酸度(TA)。用比色计(日本 Konika Minolta CR-10)测定颜色参数。其中包括亮度值(L *)、红绿色调(a *)和黄蓝色调(b *)。然后,把浆果压碎提取汁液。该果汁的SSC是用手持折射仪(数字手持口袋折射仪PAL-1,Atago,日本)测量。用氢氧化钠酸碱滴定法测定TA,终点pH为8.2,TA含量以酒石酸含量表示。剩余的浆果立即冷冻在液氮中进行代谢组学分析。
2.4 广泛靶向的代谢组学方法
2.4.1 样品制备和代谢物提取
首先,使用真空冷冻干燥机(Scientz-100F,中国宁波)对24个冷冻葡萄样品进行冷冻干燥。使用混合机(MM 400,Retsch,Shanghai,China)以30Hz 将干燥的样品研磨1.5分钟。样品的提取过程如Yang等人所详述的那样进行。具体来说,每个样品100毫克的粉末称重并溶解在1.0毫升提取液(70%甲醇水溶液)中。提取时间为24小时,提取温度为4°C。然后,将混合物以10,319 × g离心10分钟,并在UPLC-ESI-MS/MS分析之前分离和过滤上清液(SCAA-104,0.22μm孔径; ANPEL,中国上海)。
2.4.2 液相测试条件
利用HPLC: Shimpack UFLC Shimadzu cBM30a 系统, www.Shimadzu.com.cn/; MS: Applied Biosystems 6500 Q TRAP, www.appliedbiosystems.com.cn/ ,分析样品提取物。根据以前的报道,在以下 UPLC 条件下分析24个葡萄样品:柱,Agilent SB-C18(1.8 μm,2.1 mm×100mm)。流动相A组分为0.1% 甲酸和纯水。流动相B组分为0.1%甲酸和乙腈。A:B (v:v)梯度程序如下:0分钟时为95:5(v:v),9.0分钟时为5:95(v:v),10.0分钟时为5:95(v: v),11.1分钟时为95:5(v:v)和13.0分钟时为95:5(v:v)。流速保持在0.35 mL/min。将柱烤箱设置为40 °C,注射体积为4μL。出水交替连接到电喷雾三重四极线性离子阱(QTRAP)-质谱。
2.4.3 质谱条件
我们使用三重四极线性离子阱质谱仪(Q TRAP) ,AB4500 Q TRAP超高效液相色谱仪/MS/MS系统,配备有ESI Turbo离子喷雾接口,在正离子模式下工作,并由 Analyst 1.6.3软件(AB Sciex,Framingham,MA,USA)控制。根据以前的报道选择了ESI源操作参数。根据这一时期的代谢物洗脱情况监测每个时期的特定MRM转变。
2.5 统计分析
2.5.1 主成分分析和层析聚类分析
利用主成分分析和层析聚类分析初步了解24个葡萄样本之间的整体代谢差异。在无监督主成分分析之前,数据采用单位方差缩放。HCA的分析结果以热图和树状图的形式显示。PCA和HCA均采用R包热图法进行。
2.5.2 差异代谢物分析
成对比较获得的显著差异代谢物在以下条件下进行筛选:VIP ≥1和绝对 log2FC (倍数变化)≥1。VIP值从OPLS-DA结果中提取,其中还包含使用R软件包MetaboAnalystR生成的评分图和排列图。数据在OPLS-DA之前进行对数转换(log2)和均值居中。为了避免过度拟合,进行了排列测试(200种排列)。
2.5.3 KEGG注释和富集分析确定的代谢物使用
KEGG化合物数据库进行注释,然后将所有注释的代谢物映射到KEGG Pathway 数据库。将具有显著调节代谢物的途径输入到代谢物集合富集分析(MSEA)中,并使用超地理学检验的p值确定其显著性。
3 结果及分析
3.1 气象特征
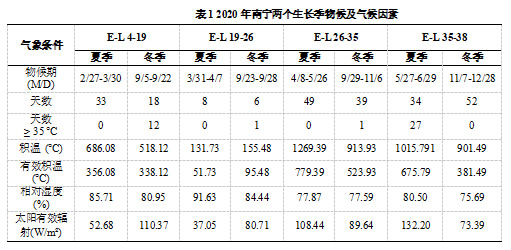
一年两收的栽培模式,充分利用南亚热带地区下半年的温光资源生产冬葡萄。表1显示了夏季和冬季葡萄生长阶段物候期和气候条件的巨大差异。夏葡萄的生长季节是从二月下旬到六月下旬,冬葡萄的生长季节是从九月上旬到十二月下旬。一般来说,夏果从E-L4(第一叶组织可见)到E-L35(变色)的时间比冬果长,尤其是从E-L4到E-L19(开花初期)。然而,从E-L35到 E-L38(收获-成熟) ,冬葡萄比夏葡萄需要18天的时间。对于夏季葡萄,有27天的高温超过35 °C,从E-L 35到E-L 38。夏葡萄的活跃积温和有效积温均高于冬葡萄,但E-L19-E-L26(帽落完全)除外。此外,相对湿度在夏季生长期较冬季生长期为更高。在第二生长季,太阳辐射强度高于第一季,从E-L4到E-L26。然而,对于第二季E-L26到E-L38,结果却是相反的。
3.2 夏葡萄和冬葡萄的理化指标
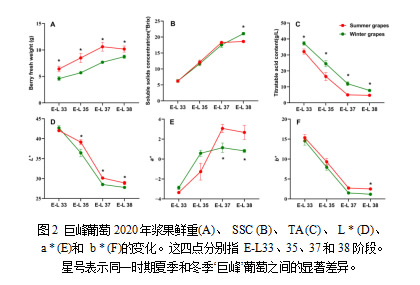
为了比较不同发育阶段夏葡萄和冬葡萄理化指标的差异,测定了不同发育阶段的浆果鲜重、SSC、TA和色泽特性。与冬季葡萄相比,夏季葡萄在四个阶段的每个阶段都显示出明显较高的浆果鲜重(图2A)。此外,与夏季葡萄相比,冬季葡萄在E-L38的SSC显著增大,但在其他阶段没有显著差异(图2B)。在整个发育阶段,冬葡萄的TA含量显著高于夏葡萄(图2C)。两种作物的葡萄的颜色属性表明L *和b *的值逐渐减少(图2D,F) ,a *值逐渐增加(图2C)。此外,夏葡萄在整个成熟期的L *值显著高于冬葡萄,这表明冬葡萄颜色较深(图2D)。另一方面,成熟后期冬葡萄的a *和b *值显著高于夏葡萄(图2E,F)。这说明冬葡萄的红色和黄色调比夏葡萄的弱。
3.3 两季葡萄的代谢产物概况
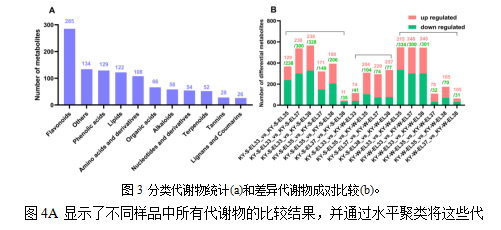
研究了四个发育阶段的代谢产物特征,特别是夏季和冬季葡萄的代谢产物差异。广泛靶向的代谢组学分析揭示了本研究中的1062种代谢物(表 S1),包括类黄酮(285),酚酸(129),脂质(122),氨基酸和衍生物(108),有机酸(66),生物碱(58),核苷酸和衍生物(54),萜烯(52),单宁(28)和木脂素和香豆素(26)(图3A)。为了澄清同一生长季节和同一发育阶段的两种作物之间的不同代谢物,计算了上调和下调的代谢物的数量,如图3B所示。对于夏季和冬季的葡萄来说,绿果期(E-L33)和成熟期(E-L35,E-l37和E-L38)的成对比较结果是相似的。具体来说,下调的代谢产物数量高于上调的代谢产物数量。就成熟期样本的成对比较而言,夏果和冬果的结果有所不同。首先,夏果的差异代谢物总量高于冬果。其次,在成熟期,成对比较上调的代谢产物数量高于下调的代谢产物数量。值得注意的是,在同一发育阶段冬果和夏果的比较中,大多数代谢产物上调。此外,三个成熟期的差异代谢产物数量均高于绿果期。
谢物分为两组。第一组代谢产物在夏、冬葡萄成熟期优先积累,第二组代谢产物在绿果期积累较多。PCA将24个样本分为三组,结果与聚类树状图一致(图4B)。结果表明,绿果期代谢产物的组成和含量与成熟期有显著差异。同时,E-L35、 E-L37和 EL38的夏果和冬果也存在显著差异。
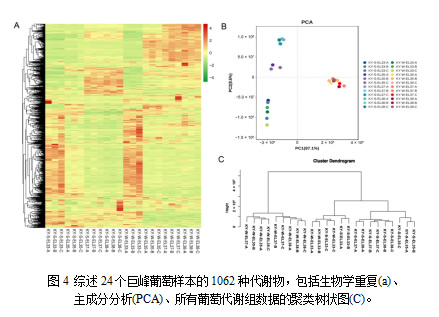
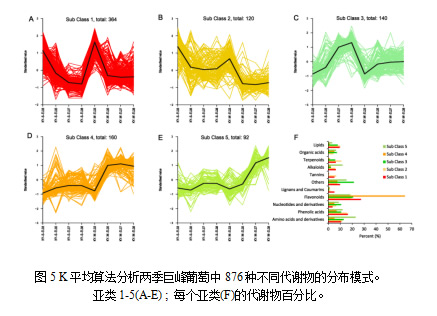
3.4 代谢物积累情况
为了更清楚地分析发育过程中代谢物的积累特征,我们对夏季和冬季葡萄进行了K平均算法分析。8个葡萄样品中的876种差异代谢物根据代谢谱差异被聚类为5个亚类(图5)。E-L33时期总共484个代谢产物在亚类1和2显示了较高的含量 (图5A,B)。E-L35、E-L37和E-L38葡萄中3、4和5亚类的392种代谢物含量较高(图5C-E)。总体而言,黄酮类化合物在每个亚类中的比例最高;特别是在亚类4中,比例高达64% (图5F)。
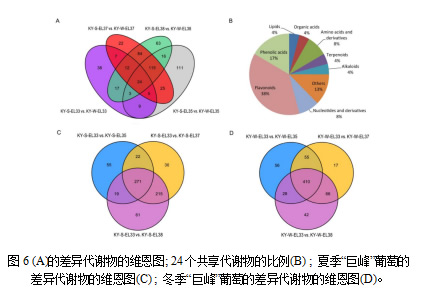
维恩图说明了不同发育阶段的差异代谢物之间的关系(图6A),分别在E-L 33,35,37和38中特异性检测到38,111,22和63种差异代谢物。这表明,特征代谢物存在于夏季或冬季葡萄的转色期 (E-L 35)。所有四个阶段共有24种不同的代谢物(表S2) ,每个类别的比例如图6B所示。具体来说,黄酮类化合物的比例最高,其次是酚酸类化合物。杨梅素-3-O-阿拉伯苷是冬葡萄中黄酮醇含量始终高于夏葡萄的一种。此外,同一生长季节的青果期和成熟期之间的成对比较结果如图6 c,d所示。值得注意的是,对于夏季和冬季葡萄,三个对照组分别共有271和410种不同的代谢物。推测冬葡萄果实发育过程中差异代谢产物主要存在于整个发育过程中。
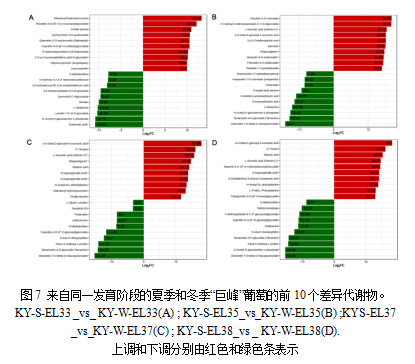
为了发现两个对照组之间存在较大差异的代谢物,我们使用Log2FC对前10种物质进行了排名,包括上调和下调。夏葡萄和冬葡萄在不同发育阶段存在明显差异。具体而言,对于KY-S-EL33_vs_KY-W-EL33具有最大Log2FC的上调和下调物质分别是双酒石酰羟基香豆素和红薯酸(图7A)。对于KY-S-EL35_vs_ KY-WEL35,具有最大Log2FC的上调和下调物质分别是patuletin-3-O-芸香苷和薯蓣蛋白-7-O-β-D-吡喃葡萄糖苷(图7B)。值得注意的是,KY-S-EL37_vs_KY-W-EL37和KY-S-EL38_vs_KY-W-EL38显示了类似的比较。例如,具有最大Log2FC的上调和下调物质是4-O- β-D-葡萄糖基-4-香豆酸和薯蓣蛋白-7-β-D-吡喃葡萄糖苷(图7C,D)。
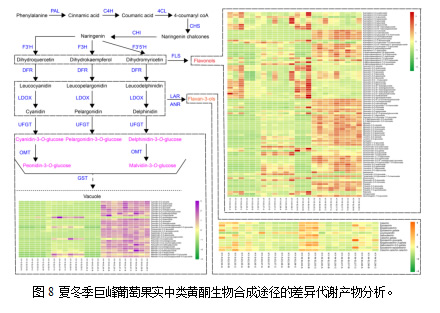
3.5 两季葡萄中黄酮类化合物的差异代谢
研究表明,黄酮类化合物是造成对照组间差异的主要原因。因此,我们重点分析了24个样品中136个差异代谢的黄酮类化合物(图8)。具体来说,这些化合物包括95种黄酮醇、12种黄酮-3-醇和29种花青素。黄酮类化合物在夏、冬葡萄成熟后期的含量高于E-L33和 E-L35,主要包括某些山奈酚、槲皮素和异鼠李素,以及所有的杨梅酮、利莫西亭和棒曲霉素。此外,这些化合物在冬葡萄中的含量高于夏葡萄。值得注意的是,夏葡萄成熟后期某些化合物的含量高于冬果,如山奈酚-4′-O-葡萄糖苷、山奈酚-3-O-芸香苷-7-O-葡萄糖苷、6-甲氧基山奈酚-3-O-葡萄糖苷、异鼠李素-3-O-阿拉伯糖苷、异鼠李素-3-O-葡萄糖苷和异鼠李素-7-O-葡萄糖苷。对于黄烷-3-醇,E-L33期的大部分化合物含量高于夏、冬葡萄的春化期和后熟期。此外,除了无色花色素外,冬果中的3-醇类物质含量高于夏果。而在夏葡萄成熟后期,无色花色素含量较高。花色苷含量在夏、冬两季葡萄发育过程中逐渐积累。除花青素 -3-O-葡萄糖苷和二甲花翠素-3-O-乙酰葡萄糖苷外,冬葡萄成熟期16个样品中大部分花青含量均高于其他样品。
4 讨论
近年来,广泛靶向的代谢组学方法已被用于检测各种果树植物的代谢物特征,如柑橘,枇杷,龙眼,猕猴桃,苹果,芒果和葡萄。采用广泛靶向的代谢组学方法获得的代谢产物数量一般在200-1000之间,这对于研究不同品种代谢产物组分的特征非常重要。2022年,在两个酿酒葡萄品种中使用广泛靶向的代谢组学方法共检测到774种代谢物,400种次级代谢物中有157种类型的黄酮类化合物。本研究在夏冬季巨峰葡萄中共检测到1062种代谢产物,其中黄酮类化合物285种,酚类化合物129种,脂类化合物122种,氨基酸及衍生物108种,有机酸66种,生物碱58种,核苷酸及衍生物54种,萜类化合物52种,单宁28种,木脂素及香豆素26种。根据上述分析,巨峰葡萄的578个次生代谢产物中含有285种黄酮类化合物。因此,黄酮类化合物在食用葡萄和酿酒葡萄的所有代谢产物中占很大比例。此外,使用传统的HPLC-MS技术,在巨峰葡萄中共检测到25种类型的花青素。用UPLC-QQQ-MS对所有样品中的37种花色苷进行了验证。
在检测到的1062个代谢产物中,有876个代谢产物在四个发育阶段不同,551个代谢产物在同一时期夏果和冬果之间不同。有趣的是,无论是发育和两季葡萄的比较表明,最主要的差异代谢物是类黄酮。葡萄中的黄酮类化合物主要由黄酮醇、黄烷-3-醇和花色苷组成,具有多样性、特异性和发育特异性的积累特征。其中,黄酮醇在果实发育的早期和成熟阶段合成并在果皮中积累,黄烷-3-醇在果实发育的早期合成并在果皮和种子中积累,花色苷在果皮中合成并积累到完全成熟。在本研究中,三种类型的黄酮在发育过程中的变化也符合上述研究,并且与我们的研究小组在苯丙烷-类黄酮合成途径中获得的两季葡萄的差异基因表达情况一致。
与酿酒葡萄相比,鲜食葡萄代谢产物的研究更多地集中在糖、酸和香气物质上。作为一种经典的鲜食红葡萄品种,巨峰葡萄的花青素含量有很多研究。但是,目前国内外对其中酚类物质,特别是黄酮类物质的组成特征的研究还很少。糖在葡萄中黄酮类化合物尤其是花青素的合成中起着重要作用,它可以促进花青素的积累。在本研究中,冬葡萄花色苷含量和可溶性固形物含量均高于夏葡萄。因此,推测夏葡萄糖分不足可能是影响花青素合成的重要因素。以前的研究发现,高温(>35°C)可以通过抑制与类黄酮代谢相关的基因的表达,影响酶活性并降低ABA 和类黄酮前体(苯丙氨酸)的含量来促进花色苷降解。与花青素类似,高温抑制了黄烷-3-醇的关键基因ANR和LAR的表达。对于一年两熟的栽培系统来说,夏季生长季的极端高温天数是冬季生长季的两倍。这无疑是夏葡萄中黄酮类化合物含量较低的一个重要原因。
UVB和UVC的辐射可以促进葡萄中黄酮醇和黄烷-3-醇的积累。作为光信号成分的CRY2和HY5/HYHs,作为转录因子的MYBA1,以及作为关键结构基因的LAR,ANR和FLS,负责相应黄酮类化合物的光响应生物合成。冬葡萄早期的光照强度高于夏葡萄,这可能是冬葡萄中黄酮醇和黄烷-3-醇积累较多的重要原因。此外,水分不仅影响葡萄的物理性质,而且对葡萄内部代谢产物也有重要影响。在我们的研究中,冬季葡萄在生长季节的相对湿度低于夏季葡萄,而较干燥的环境也可能导致较高的类黄酮合成。较低的湿度可以通过降低浆果鲜重或直接上调黄酮合成途径相关基因的表达来促进黄酮类化合物的积累。
5 结论
本研究采用广泛靶向的代谢组学方法,系统评价了两季巨峰葡萄不同生育期代谢产物的差异。共检测到1062种代谢产物,其中黄酮类化合物285种。通过聚类分析和差异代谢产物分布分析,确定黄酮类化合物是葡萄在两种作物之间和四个发育阶段之间分化代谢产物的最重要组成部分。总体而言,不同发育阶段的差异大于同一发育阶段夏葡萄和冬葡萄的差异。在四个发育阶段共鉴定出876个差异代谢产物,在同一时期夏季和冬季葡萄中共鉴定出551个差异代谢产物。此外,16个代谢产物组成和含量在绿果期与成熟期有显著差异。但是,成熟期的差异代谢产物数量高于绿果期。夏冬葡萄生长季气候条件的不同是造成两季葡萄类黄酮代谢产物差异的主要原因。具体来说,夏季葡萄生长季节的极端高温日数较多,相对湿度较高,导致夏季葡萄的类黄酮含量较冬季葡萄为低。此外,冬葡萄中大量黄酮醇和黄烷-3-醇的积累可能与早期果实发育过程中较强的光照强度有关。杨梅素-3-O-阿拉伯苷在冬葡萄发育各阶段均高于夏葡萄。值得注意的是,大多数黄酮类化合物在冬葡萄中含量较高,但在夏葡萄中也有一些含量较高,如山奈酚-4′-O-葡萄糖苷、无色花色素和花色素-3-O-葡萄糖苷。综上所述,本研究采用基于UPLC-QQQ-MS的代谢组学方法检测了巨峰葡萄的主要和次要代谢产物。此外,研究结果还揭示了造成夏季和冬季葡萄代谢产物差异的气候因素。未来的研究可能集中在气候因素调节品质相关代谢产物的机制方面统。另外,通过对具体栽培技术的研究,希望能够克服夏冬两季葡萄栽培中的一些缺陷。