栽培生理
摘 要:为探索两个葡萄品种在怀涿盆地的光合生产情况,以5年生‘阳光玫瑰’和‘南太湖特早’葡萄为材料,测定其在草炭、牛粪、玉米秸秆、菌棒渣等4种基质处理下的光响应曲线、CO2响应曲线与光合速率日进程等光合特性,并测定果实品质。结果表明:‘南太湖特早’在草炭与牛粪处理下具有较高的光合能力、表观量子效率以及羧化效率,草炭处理光补偿点更低,牛粪处理果实糖酸平衡表现更佳,果实品质较好;‘阳光玫瑰’在玉米秸秆和草炭处理下表现出较高的光合能力与羧化效率,表明其对‘阳光玫瑰’的光合作用有利,其中玉米秸秆处理下的糖酸比、花青素最佳,有利于提高‘阳光玫瑰’的果实品质。
关键词:葡萄;光合特性;基质;果实品质
我国是葡萄栽培大国,葡萄栽培面积广、栽培历史悠久[1]。怀来县地处怀涿盆地,自然条件优越,被誉为中国葡萄之乡[2],其主栽的鲜食葡萄以‘龙眼’‘白马奶’等传统品种为主[3],品种结构较为单一。为丰富怀涿盆地鲜食葡萄品种,当地引进了‘阳光玫瑰’‘南太湖特早’等新品种。有研究表明怀来大部分葡萄园处在冲积平原,土壤中黏粒与有机质含量少[7],导致肥水流失、利用率低等问题。因此,许多研究尝试使用菌棒渣、草炭等农业副产物或有机基质来改良土壤。刘双双[4]研究表明有机基质栽培可以提高‘巨峰’葡萄果实的外观及内在品质,并使其成熟期提前;秦玲等[5]认为草炭基质可以显著改善葡萄根系,促进地上部分和根系的平衡生长。李博[6]研究表明以蚯蚓粪为主的基质可以提高多种葡萄扦插苗的生长量及其果实品质。但目前关于不同基质栽培对葡萄光合特性的影响鲜有报道。
光合作用是植物生长发育的基础,葡萄叶片的光合特性和光合能力直接影响葡萄的果实品质和产量。本试验以5年生‘阳光玫瑰’和‘南太湖特早’葡萄为试材,对葡萄在不同基质处理下的光响应曲线、CO2响应曲线与光合速率日进程等光合特性进行研究对比,探索两个葡萄品种在怀涿盆地的光合生产情况,为其高质量生产提供理论依据。
1. 材料与方法
1.1 试验地概况
试验地点位于河北省张家口市怀来县城投农业开发有限公司所属的设施葡萄栽培园区(40°20’N,115°33’E),平均海拔535 m,年平均降水量396 mm,年平均温度9.1 ℃,年均日照时数3027 h,无霜期149 d(数据来源:怀来县人民政府)。
1.2 试验材料
试验品种为5年生‘南太湖特早’与‘阳光玫瑰’葡萄,均为嫁接苗,砧木为SO4,设施栽培,株行距为1.5 m×3 m,东西行向,主干高约2 m,厂型架,灌溉方式为滴灌与漫灌结合。
于2022、2023年春季树体萌发前,在栽植沟内铺施厚10 cm、宽120 cm的草炭、牛粪、玉米秸秆、菌棒渣4种不同基质,其中草炭(进口)、牛粪(当地牧场)、菌棒渣(张北)为腐熟后的材料,玉米秸秆为(试验地附近所产)直接打碎后的材料,每种基质铺施两沟,旋耕机翻土将基质和园土混匀,无基质铺施的两个栽植沟作为对照处理。两品种均在秋季落叶期开40 cm宽沟施入等量羊粪(厚度10 cm)作为基肥。具体如表1:
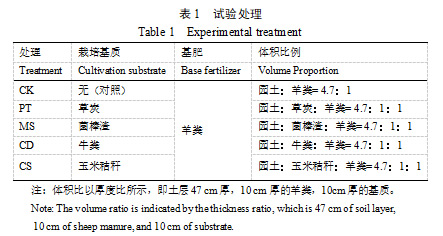
1.3 试验方法
1.3.1 葡萄叶片光合特性的测定
每个处理选取12个长势基本一致且具代表性的结果母枝进行标记,测定其果穗对生叶片的光响应曲线、CO2响应曲线及光合日进程,试验仪器为德国WALZ公司生产的GFS-3000型便携式光合仪,探头型号为标准探头3010-S,光源为3041-L,测定时间为2024年8月21日-31日。
Pn-PAR光响应曲线的测定:选择晴朗无云的上午(9:00—11:00)测定叶片在不同光合有效辐射(PAR,photo-synthetically active radiation)下的净光合速率(Pn, Photosynthetic rate),3次重复。将仪器开机,待光合系统预热完成后设定叶室内各项参数:叶室温度(Tcuv)26℃、参比室CO2浓度(CO2abs)为400 mg·kg-1、参比室H2O浓度(H2Oabs)为绝对值18000 mg·kg-1,叶室顶部光源的光合有效辐射(PARtop)梯度为0、50、80、100、200、400、600、800、1000、1200、1400、1600、1800 μmol·m-2s-1。
Pn-CO2响应曲线的测定:选择晴天的上午(9:00-11:00)进行测定,重复3次,设定光合测定系统各项工作参数:叶室PARtop为1600 μmol·m-2s-1,温度为26℃,H2O浓度为绝对值18000 mg·kg-1,CO2浓度梯度为50、100、200、400、600、800、1000、1200、1400、1600、1800 mg·kg-1。
光合日变化特性的测定:选择无风的晴天,从早上7:00至下午6:00,每隔1h进行一次光合特性的测定,测定指标为净光合速率(Pn),胞间CO2浓度(Intercellular CO2 concentration, ci),蒸腾速率(Evaporation rate, E),重复3次。
1.3.2 葡萄果实品质的测定
2024年,在两个品种果实达到商品果标准(可溶性固形物≥18%)后开始采样,每个处理随机挑选5串具有代表性的果穗,在果实上、中、下部位各取3个果粒,挤压出果汁,使用手持数显折光仪PAL-1测定可溶性固形物(SSC),采用酸碱滴定法测定可滴定酸含量(TA),并计算果实固酸比(SSC/TA,%),每串果穗作为1个重复,重复5次。
1.4 数据分析
采用SPSS 26.0软件进行方差分析,采用光合计算软件进行光合曲线拟合以及光合参数计算,具体计算公式与方法参考叶子飘等[10-11]的回归模型。
2. 结果与分析
2.1 不同基质处理下葡萄的光响应曲线特性
不同处理下两个葡萄品种的光响应曲线较为相似,其净光合速率(Pn)均随着光合有效辐射(PAR)的增加而逐渐提升,提升幅度由大变小,最后逐渐趋于饱和(图1)。PAR在0~400 μmol·m-2s-1区间时,各处理净光合速率随光强的增加提升幅度最大,而在PAR增加到1000 μmol·m-2s-1后,各处理的净光合速率均已趋近最大值。在各PAR下,两葡萄品种在CD、CS和PT处理下的净光合速率与对照相比均有提高,MS处理则低于对照。
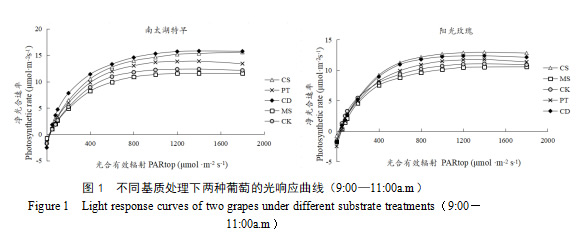
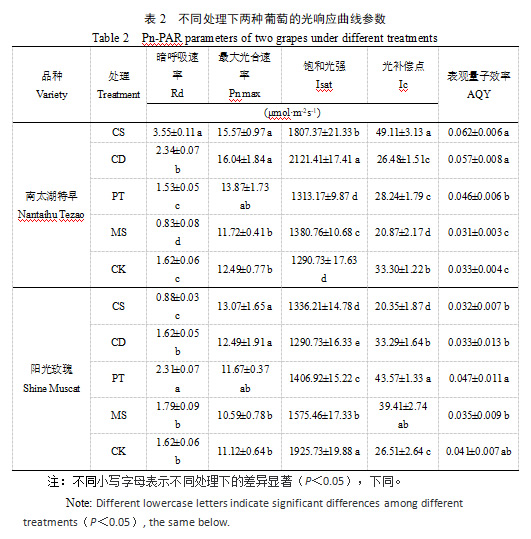
将光响应数据代入光合模型[11]进行曲线拟合与计算得出的光响应曲线参数,如表2所示。不同基质处理对两葡萄品种的光合参数具有显著影响。对于‘南太湖特早’,CS处理的暗呼吸速率(Rd)较CK显著提高了119%,同时表现出较高的最大光合速率(Pn max)和表观量子效率(AQY),说明该基质促进光合作用的同时也增加了呼吸消耗。CD处理的Pn max和饱和光强(Isat)最高,且光补偿点(Ic)较低,表明该基质能提高光能利用效率,使植株在较低光强下即可维持光合作用。对于‘阳光玫瑰’,PT处理的AQY较高,但Rd和Ic也较高,表明该基质可能在促进光合作用的同时增加了呼吸消耗;对照处理的Isat显著高于其他处理,说明自然土壤环境更有利于‘阳光玫瑰’适应高光强条件。
2.2 不同基质处理下葡萄的CO2响应曲线特性
如图2所示,两个葡萄品种的CO2响应曲线与其Pn-PAR曲线较为相似,净光合速率会随胞间CO2浓度的提高而提高,最后趋近饱和。在Ci小于600 μmol·mol-1时,两品种的Pn值提升最快。其中‘阳光玫瑰’的CK、MS处理在CO2饱和之后,继续增加CO2浓度,净光合速率会出现明显的下降趋势。
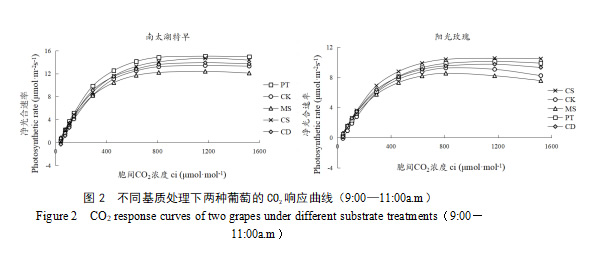
将CO2响应数据带入光合模型[11]进行分析后可得出表3所示的各项光合参数。由表3可知,MS处理下的‘南太湖特早’光呼吸速率(Rp)较CK显著降低了36.2%,‘阳光玫瑰’Rp对比CK显著降幅52.4%,表明MS处理能降低葡萄呼吸消耗,但同时光合能力与羧化效率也较低。‘南太湖特早’在PT处理下光合能力(Pn max)最高,较CK提高11.9%,而‘阳光玫瑰’在CS处理下Pn max提升了15.0%,同时PT处理还使两品种CO2饱和点(Cisat)提高16.4%~24.4%,羧化效率(CE)平均增加20.9%。CD处理显著提升了‘南太湖特早’的羧化效率(较CK增加33.3%),同时导致光呼吸显著增强(Rp较CK增加62.4%)。
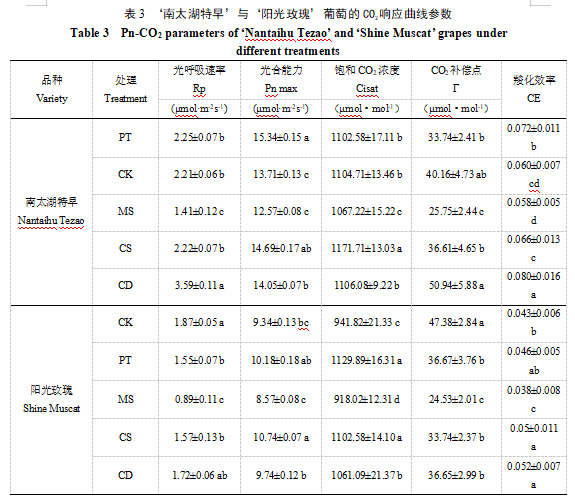
2.3 不同基质处理下葡萄的光合日变化特征
两品种的净光合速率均呈现明显的双峰曲线,分别在10:00—11:00和13:00—14:00出现两个峰值,且第二个峰值小于第一个(图4)。正午(11:00—13:00)期间,光合速率显著下降,表现出典型的“光合午休”现象[8]。
‘南太湖特早’的CD处理整体光合速率较高,且在上午10:00—11:00时表现出最高的光合速率,峰值接近14 μmol·m⁻²s⁻¹;CS与PT处理的光合速率次之,且在13:00—14:00的第二个峰值时,表现出较好的光合作用恢复能力;MS处理光合速率最低,尤其在上午11:00时明显低于其他处理,且光合速率恢复较慢。‘阳光玫瑰’的PT与CK处理在上午达到较高的光合速率峰值,表现出更强的光合能力;CS和CD次之,表现也较为稳定。MS的光合速率整体偏低,尤其在高光时段明显受抑制。
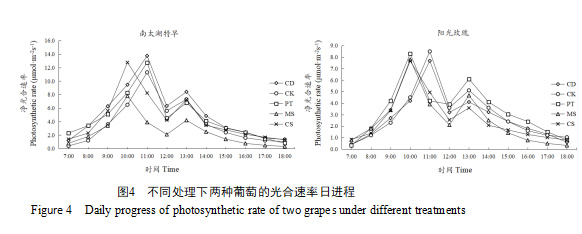
如图5所示,‘南太湖特早’与‘阳光玫瑰’在不同处理下的蒸腾速率变化趋势与其光合速率变化趋势较为相似,也呈现出双峰曲线,分别出现在10:00—11:00和13:00—14:00。‘南太湖特早’的PT处理在上午出现最高峰值,为3.5 mmol·m-2s-1;CS处理的在上午峰值也较高,仅次于PT处理,且表现出较强的恢复能力,在下午达到的峰值最高,为2.19 mmol·m-2s-1。对于‘阳光玫瑰’,PT和CD处理的蒸腾速率在上午表现出较高的峰值,其值分别为3.18、3.13 mmol·m-2s-1。
2.4 不同基质处理下葡萄果实品质
不同有机基质处理对两种葡萄果实品质调控作用显著(表4)。‘南太湖特早’在CS处理下可溶性固形物含量最高,较CK提高13.9%,但可滴定酸较CK显著升高15.7%,导致固酸比显著最低;而MS与CD处理显著降低了酸含量(较CK降31.4%~22.9%),固酸比显著提升36.0%~36.3%,有效改善了糖酸协调性。‘阳光玫瑰’ PT、CD和CS处理下可溶性固形物均高于20%(较CK提高16.1%~21.6%),其中CS处理固酸比达70.47(较CK提高11.2%),实现糖酸同步优化;而MS处理可滴定酸显著升高(较CK增20.7%),导致固酸比最低。
3. 讨论
光响应曲线是判断植物对光能利用效率的重要依据之一,通过分析光响应曲线,可以得到植物的表观量子效率以及光饱和点等相关参数,以此了解植物的光合作用能力[12]。表观量子效率(AQY)反映了植物光合作用中光能转化为化学能的效率,其理论最大值约为0.125,在实际自然条件下,长势良好的作物AQY值通常在0.04~0.07之间[16]。本试验中,‘南太湖特早’和‘阳光玫瑰’两种葡萄在草炭(PT)处理下的AQY值均高于0.04和对照处理,这表明草炭基质有利于提高葡萄的光合能力,其原因可能是草炭基质改良砂土后,增加了土壤的孔隙度和田间持水量,从而提高葡萄的光合速率[5]。光补偿点反映了植物在弱光下的光合能力,光补偿点越低,植物利用弱光的能力越强[14];饱和光强则反映了植物在强光条件下的光合能力。饱和光强越高,说明植物利用强光的能力越强,在强光照射下越不易发生光抑制[15]。本试验发现‘南太湖特早’的玉米秸秆(CS)和牛粪(CD)处理AQY值比PT处理更高,但同时呼吸消耗显著增加,CS处理的光补偿点显著提高,CD处理的饱和光强显著提高,说明CS和CD在光照强度较高的环境才能发挥优势。相比而言PT处理的光补偿点和饱和光强较低,说明其在设施内弱光条件下表现良好。
CO2是植物光合作用的重要原料,是限制植物光合作用的主要因子[20-22],通过CO2响应曲线可以确定植物的光呼吸速率、CO2补偿点等光合参数,同时可以估算出植物的最大羧化效率[17]。本试验发现‘南太湖特早’在PT和CD处理下具有较高的光合能力和羧化效率,但CD处理同时使其光呼吸速率显著提高,增加了其呼吸消耗,需谨慎使用。 而‘阳光玫瑰’葡萄在CS处理下的光合能力最高,同时羧化效率显著提高,呼吸消耗显著下降,有利于提升该品种的光合潜力。在本试验中两个葡萄品种在不同基质处理下的光合速率变化均呈现典型的双峰曲线,在中午时段表现出明显的“光合午休”现象,这与前人的研究结果一致[18-19]。
光合作用是植物将光能转化为化学能的过程,是葡萄糖分积累的关键。本试验中PT、CD与CS处理均显著提高了‘南太湖特早’与‘阳光玫瑰’的可溶性固形物含量,而固酸比方面‘南太湖特早’的CD与菌棒(MS)处理及‘阳光玫瑰’的CS处理有显著提高,这是由于CS处理在提升两品种糖积累的同时使‘南太湖特早’的酸度增加。CS可以促进土壤养分转化,改善土壤物理性质,保蓄土壤水分,平衡土温,增加田间CO2浓度,提高产量和品质,但秸秆直接还田携带大量病菌虫卵,对改土效果,有机质积累和作物生长有一定影响[23]。本试验中玉米秸秆并未经过充分降解,这可能是影响果实酸度的因素之一。MS处理显著改善了‘南太湖特早’的糖酸平衡,但同时导致‘阳光玫瑰’酸度过高。菌棒渣还田可以降低土壤pH,同时增加土壤EC值,提高土壤有机质含量[24],但如果使用不当也容易造成烧苗,增加表层土壤盐渍化的风险。相对而言草炭与牛粪基质对‘阳光玫瑰’果实品质提升效果稳定。
4. 结论
综合分析认为,两个供试品种的净光合速率呈现双峰曲线,存在光合午休现象。处理间比较后发现,‘南太湖特早’在草炭与牛粪处理下具有较高的光合能力、表观量子效率以及羧化效率,其中草炭处理的光补偿点和CO2补偿点更低,且在光合日进程的午休现象过后显现出较强的恢复能力,光合功能优秀;牛粪处理果实品质较高,糖酸比最佳,果实品质较好。两个处理均提高了果实糖分积累,适合提高葡萄的果实品质。‘阳光玫瑰’在玉米秸秆处理下表现出较高的光合能力与羧化效率,且糖酸比最佳,有利于提高‘阳光玫瑰’的果实品质。