鲜食葡萄品种改良
1 材料与方法
1.1 植物材料
本研究材料为两个欧亚葡萄(Vitis vinifera)栽培品种‘意大利’和‘塔米娜’及其F1杂交后代。‘意大利’是意大利Pirovane通过‘比坎’与‘玫瑰香’杂交选育的晚熟鲜食葡萄品种;‘塔米娜’为其罗马尼亚对应品种,具有完全相同的亲本来源。两个品种均具有显著的麝香/花香特性,种植于北京市农林科学院林业果树研究所(北纬39°58′,东经116°13′)。供试植株为五年生自根苗,采用水平龙干形棚架系统配合单臂叶幕整形方式栽培,行距2.5米,株距0.75米,采用常规栽培管理措施。
1.2 群体构建
杂交群体于2010年通过人工控制杂交建立,以‘意大利’为母本、‘塔米娜’为父本,对30个花序进行人工授粉后共获得535粒种子。经种子提取与层积处理后,于2011年播种获得200余株实生苗,随后定植于试验果园。2017年采集153株子代植株样品,2018年采集135株——样本量减少是由于2018年成熟期异常降雨导致的果实腐烂。分析样品采集时均衡考虑果穗阴阳面(遮光/向阳)与穗部位置(肩部/中部/尖端)的代表性,每个样本随机选取100粒成熟浆果,设置三个生物学重复,样品经液氮速冻后于-80°C超低温保存。
1.3 单萜类物质的顶空固相微萃取-气相色谱-质谱(HS-SPME-GC-MS)提取与分析
游离态单萜提取参照文献方法(Liu et al., 2022),采用安捷伦7890气相色谱-5975B质谱联用仪(安捷伦科技有限公司,美国)进行分析。色谱分离使用HP-Innowax毛细管柱(60 m × 0.25 mm × 0.25 μm;J&W Scientific),高纯氦气作为载气,流速1.0 mL/min(Liu et al., 2018)。通过全自动固相微萃取(SPME)系统进样,采用不分流模式,250°C热解吸8分钟;程序升温初始50°C(保持1分钟),以3°C/分钟速率升至220°C(保持5分钟)。质谱采用电子轰击(EI)离子源(70 eV),离子源与传输线温度分别设定为230°C和280°C,质量扫描范围m/z 29-350。
1.4 单萜类物质的定性与定量分析
保留指数(RI)和质谱信息通过自动质谱解卷积与鉴定系统(AMDIS)计算,化合物鉴定通过质谱分析结果与NIST11数据库匹配完成,并结合标准保留指数进行验证。葡萄萜烯类化合物定量分析采用标准曲线法(Liu et al., 2018; Wang et al. 2018):基于葡萄汁糖酸含量配制含7 g/L酒石酸和200 g/L葡萄糖的1 L模拟葡萄溶液,用1 mol/L NaOH溶液调节pH至3.4。各标准品先用色谱纯乙醇溶解,混合后使用模拟基质溶液梯度稀释至15个浓度水平。对于缺乏标准曲线的单萜化合物,采用碳原子数与化学结构相似化合物的标准曲线进行定量。
1.5 统计分析
基于子代中单萜化合物的有无状况计算分离性状的表型比例。χ²检验采用公式 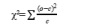
计算(o为观测值,c为理论值),并对多重检验进行Bonferroni校正。使用SPSS 27.0(SPSS公司,美国)中的K-S检验判定单萜数据是否符合正态分布或泊松分布。通过R软件绘制箱线图展示单萜浓度范围和中位数,并检测异常值:箱体上下横线分别表示样本中单萜数据的上下四分位数。采用R语言(lme4包)进行方差组分分析(ANOVA),以基因型为随机效应估算广义遗传力(H² = σ²G/(σ²G + σ²e))。相关性分析使用SPSS 27.0完成,相关性热图通过ChiPlot(https://www.chiplot.online/)绘制(Abdel-Sattar et al., 2022)。
2 结果
2.1 亲代和子代的单萜组成和含量
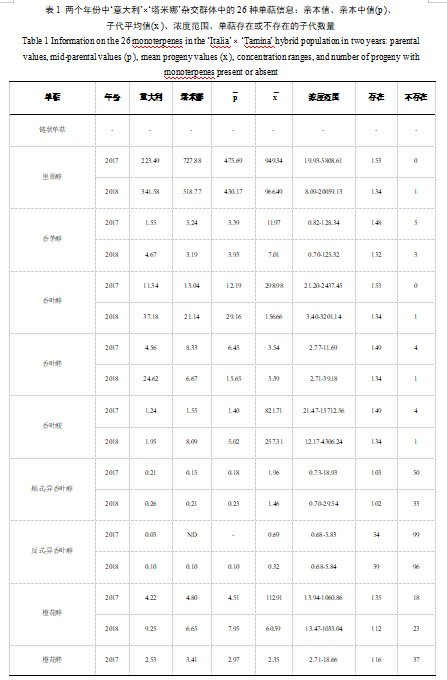
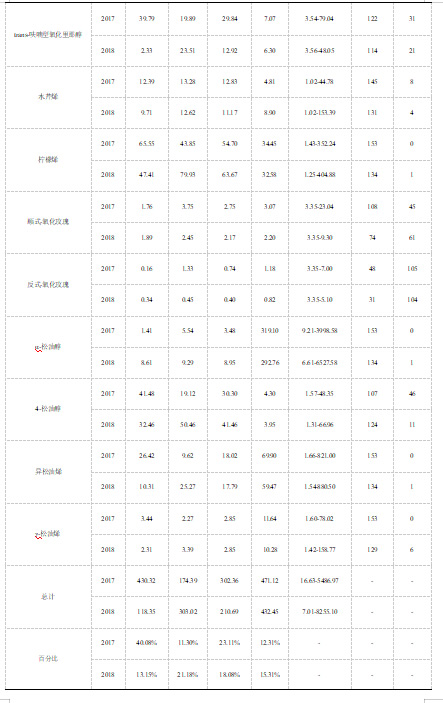
对于欧亚种亲代和其杂交的F1代葡萄群体,本研究采用顶空固相微萃取-气质联用法检测游离态单萜物质。通过连续两年的研究,我们在‘意大利’亲本中共检测到25种挥发性单萜物质,在‘塔米娜’亲本中共检测到24种单萜物质(表1)。与亲本系相比,子代中的挥发性单萜还包括trans-异香叶醇和吡喃型氧化里那醇。其中,trans-异香叶醇在‘塔米娜’亲本中未被检测到,而吡喃型氧化里那醇在两个亲本系中均未检测到。在本研究中,发现‘意大利’亲本的总挥发性单萜含量小于‘塔米娜’亲本,前者的总单萜含量为900.08-1073.73μg/kg,而后者的总单萜含量值为1430.55-1542.81μg/kg(表1)。
研究共检测到两种类型的单萜,分别为链状单萜和环状单萜(表1)。其中在‘意大利’亲本中检测到的主要挥发性单萜为链状单萜,该类型单萜的总含量在两个年份中分别为643.41μg/kg和781.73μg/kg,平均占研究期间总单萜含量的73.39%;环状单萜在该亲本的总单萜中占比相对较小,占比值为26.61%,两个年份中总环状单萜含量分别为430.32μg/kg和 118.35μg/kg。同样地,在‘塔米娜’亲本中,我们发现链状单萜仍然为主要的挥发性单萜,占研究期间总单萜含量的83.76%,而环状单萜仅占据16.24%。里那醇是含量最多的单萜,无论是在‘意大利’还是‘塔米娜’亲本中(表1)。在‘意大利’亲本中,里那醇含量值分别为223.49μg/kg和341.58μg/kg,占据总单萜含量的20.81%-37.95%。相比之下,里那醇在‘塔米娜’亲本中占据总单萜含量的36.265-47.18%,含量值分别为727.88μg/kg和518.77μg/kg,该结果高于‘意大利’亲本中的里那醇含量值。
我们在杂交子代中共检测到26种单萜物质,并利用标准曲线分别对它们进行定量。在两个年份中,总挥发性单萜的平均含量显著高于亲本中总单萜的平均含量。子代中链状单萜总含量较多,占总单萜含量的86.19%,在两个年份样本中的总链状单萜平均含量值分别为3357.16μg/kg和2392.30μg/kg。总环状单萜在两个年份中的浓度范围分别为16.63-5486.97μg/kg和7.01-8255.10μg/kg,平均含量值为451.79μg/kg,占总单萜含量的13.81%。与亲本结果相似,子代中的平均含量最高的单萜物质为里那醇,平均含量值呈现为957.92μg/kg。香叶酸的平均含量仅次于里那醇,表现为539.51μg/kg,占据子代样本中总单萜含量的15.29%。除此之外,β-月桂烯、β-cis-罗勒烯和α-松油醇的平均含量同样较高,这些物质共同构成了欧亚种杂交葡萄子代中主要的单萜组分。
此外,我们观察了26种单萜物质在全部杂交子代中的分离情况。其中,cis-异香叶醇、橙花醛、吡喃型氧化里那醇和cis-氧化玫瑰这四种单萜物质的分离情况,表明这些单萜的存在或不存在极大程度上符合孟德尔遗传定律。因此,分离模式一般符合后代中3:1或1:1的比例,但2018年橙花醛和2017年cis-氧化玫瑰分离模式的比例值明显例外(表2)。随后,通过χ²试验验证了单萜分离比例的准确性(表2)。
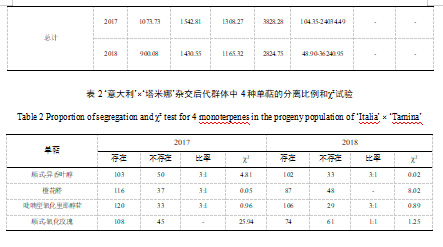
2.2 单萜物质的遗传差异分析
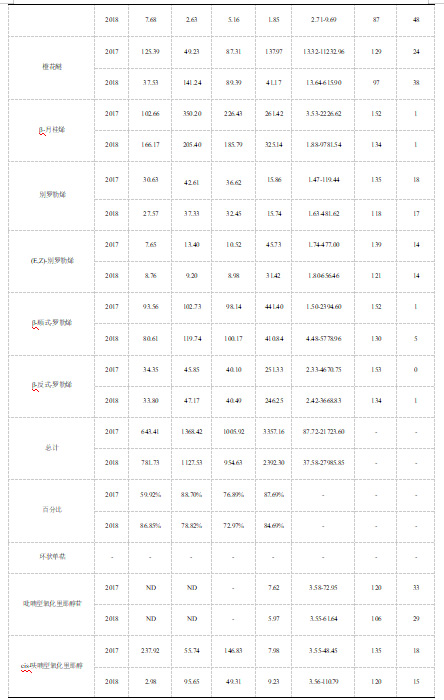
研究发现,子代中的总单萜含量平均值在两个年份中分别显示为3828.28μg/kg和2824.75μg/kg,结果显著高于亲本中的总单萜平均值1308.27μg/kg和1165.32μg/kg(表1)。在两年的研究过程中,大多数单萜物质在子代中的平均含量显著高于双亲。与双亲葡萄样本相比,里那醇在子代中的含量平均值高于双亲,其含量在子代和亲本中分别呈现为957.52μg/kg和452.93μg/kg。值得注意的是,一些单萜在子代中的含量甚至是亲本的几百倍,香叶酸在子代中的平均浓度为539.51μg/kg,是该物质在亲本中平均浓度(3.21μg/kg)的约168倍(表1)。除此之外,也有少数单萜在子代中的平均含量低于双亲。例如,子代中香叶醛的含量值低于‘意大利’和‘塔米娜’样本,该物质的平均含量在子代中呈现为3.47μg/kg,在双亲中分别呈现为14.59μg/kg和7.5μg/kg。
研究结果显示,香叶醛、橙花醛和cis-氧化玫瑰的变异系数在0.1-1.0的范围内,被认为属于中度变异;F1群体中其他单萜的变异系数均高于1.0,被认为是高度变异(表3)。检测发现,大多数单萜在分布图中浓度最小值到最大值区域的分布频率逐渐降低,表明多数单萜在杂交子代中呈现泊松分布,少数呈现正态分布(表3)。3种单萜在子代中的含量较高,分别为里那醇、香叶酸和β-cis-罗勒烯,它们均呈现泊松分布;部分单萜在杂种后代中表现出相对较低的含量,如trans-异香叶醇和trans-氧化玫瑰,它们同样呈现泊松分布。此外,两种单萜的浓度分布在不同年份中呈现不同的情况。2017年,香叶醛和橙花醛属于正态分布;而在2018年样本中两种单萜均呈现泊松分布(表3)。
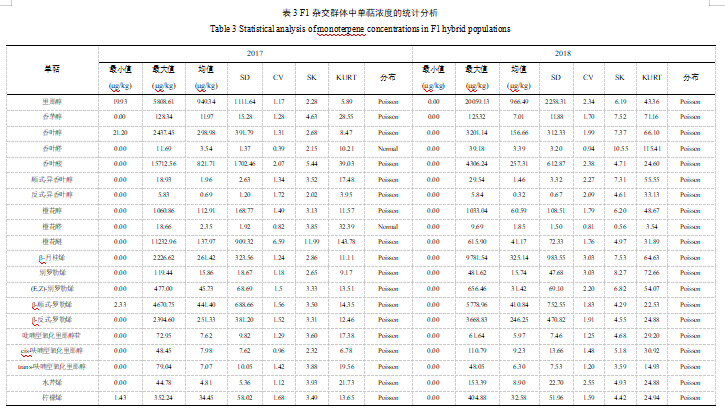
2.3 单萜物质的基因型变异和遗传
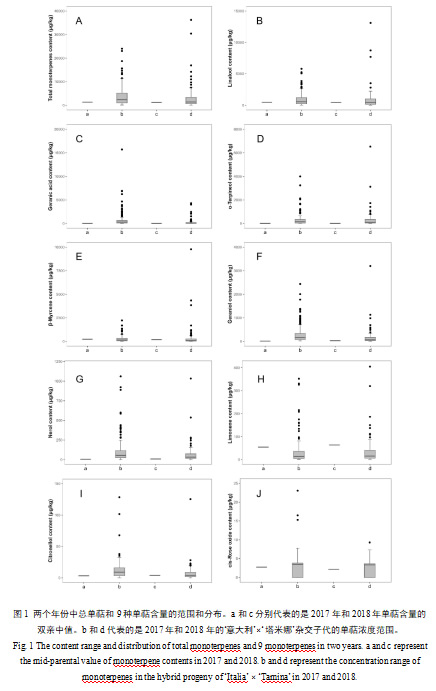
研究发现,杂交子代中的总单萜含量显著高于双亲,并呈现广泛的连续变异。2018年,总单萜的双亲中值和子代中总单萜的中位数值均低于2017年,说明年份对总单萜的积累具有较大影响(图1a)。同时,子代中总单萜的浓度分布值在两个年份中具有显著差异,2018年时总单萜的含量范围较2017年更为广泛,显示为48.90-36240.95μg/kg(图1a)。
具体来看,一些单萜物质在子代中连续变异,表现出显著的数量性状遗传(图1B-J)。里那醇在子代中的平均含量值较高,含量范围最为广泛,2017年和2018年该物质浓度范围分别呈现为19.93-5808.61μg/kg和8.09-20059.13μg/kg(图1B)。香叶酸在子代中的平均含量值次之,其分布情况同样表现出数量性状遗传(图1C);该物质在2017年的含量范围(21.47-15712.56μg/kg)显著高于2018年含量范围(12.17-4306.24μg/kg)。α-松油醇在子代中的总单萜含量中占比9.35%,该物质在子代的环状单萜中含量较高,占据总环状单萜含量的67.72%。该物质在两个年份的子代中的浓度范围分别为9.21-3998.58μg/kg和6.61-6527.58μg/kg,呈现明显的连续变异(图1D)。β-月桂烯、香叶醇、橙花醇和香茅醇的分布情况展示了其浓度分布明显趋于低值(图1E-G,图I)。此外,柠檬烯在亲本中的平均浓度值和中位数值高于其在子代中的结果,子代中柠檬烯的浓度分布呈现连续变异(图1H)。cis-氧化玫瑰的含量值最低,该物质符合孟德尔遗传定律分离,在子代中表现出数量性状遗传(图1J)。并且,cis-氧化玫瑰在双亲和子代中的平均含量差异较小,分别呈现为2.46μg/kg和2.64μg/kg。
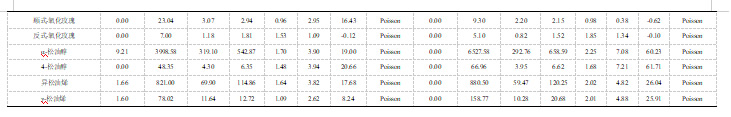
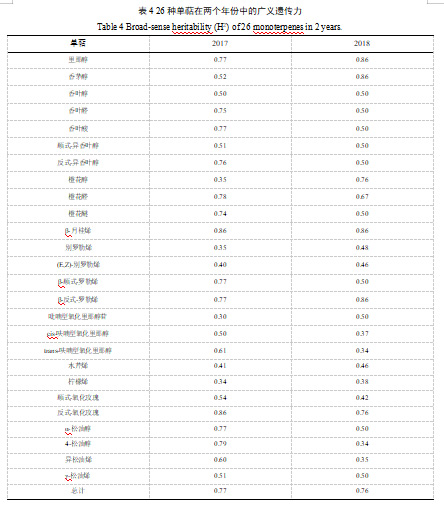
此外,我们对26种单萜进行了广义遗传力(H2)分析。研究发现,β-月桂烯含量的H2值最高,在两个年份中均呈现为0.86(表4)。别罗勒烯、(E,Z)-别罗勒烯、吡喃型氧化里那醇、cis-呋喃型氧化里那醇、水芹烯和柠檬烯这些单萜物质含量的H2值较低,它们在两个年份中的数值均不超过0.5(表4)。
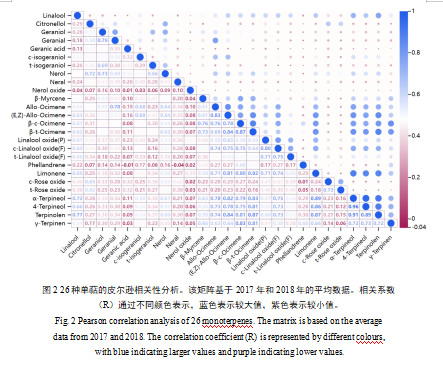
2.4 单萜物质的相关性分析
我们对26种单萜物质进行了相关性分析。总体来看,大多数单萜之间呈现显著的正相关关系,个别单萜(香叶酸和橙花醛)与水芹烯呈现负相关关系。我们发现,别罗勒烯、(E,Z)-别罗勒烯、β-cis-罗勒烯、β-trans-罗勒烯、柠檬烯、α-松油醇、4-松油醇、异松油烯和γ-松油烯与其他单萜物质的相关性较强;而香叶酸、橙花醛、橙花醚和水芹烯与其他单萜的相关性较弱(图2)。α-松油醇与4-松油醇和异松油烯的相关系数最高,分别为0.963和0.91(P<0.01);水芹烯与cis-氧化玫瑰的相关系数最低,为0.012。值得注意的是,来自相同生物合成途径的单萜烯之间呈正相关,例如橙花醛与香叶醛。此外,4种链状单萜:别罗勒烯、(E,Z)-别罗勒烯、β-cis-罗勒烯、β-trans-罗勒烯与4种环状单萜:α-松油醇、4-松油醇、异松油烯和γ-松油烯的相关性较高,相关系数值均高于0.6(图2)。