土壤与产地环境污染管控与修复
摘要:镉对葡萄的生长、发育和品质都会造成严重影响,果园受到的低浓度镉污染问题不容忽视,砧木嫁接能够提高葡萄对镉胁迫的适应能力,但关于镉胁迫下砧木嫁接后镉胁迫对葡萄的转录变化机制,尚未见报道。本研究以三种不同砧木嫁接的阳光玫瑰葡萄的二年生苗为试材,采用非靶向代谢组技术,研究了镉胁迫下三种砧穗组合叶片的代谢产物的变化。分析表明,SM/SO4组合特有的差异代谢物显著富集在氨基酸代谢、ABC转运蛋白、碳水化合物代谢通路;SM/5BB组合特有的差异代谢物显著富集在ABC转运蛋白、乙醚脂质代谢等通路;SM/3309C组和特有的差异代谢物显著富集在磷酸戊糖途径、甘油磷脂代谢等通路。同时响应镉胁迫的所有差异代谢物显著富集在氨基酸代谢、碳水化合物代谢、ABC转运蛋白、核苷酸代谢、脂类代谢、其他次级代谢物的生物合成等通路。综上诉述,镉对高等植物暂无有益作用,三种砧穗组合在低镉胁迫后代谢组层面存在着差异,代谢组WGCNA分析发现了一些响应低镉胁迫的关键基因,以及响应低镉胁迫的重要代谢通路。上述发现有助于了解嫁接阳光玫瑰葡萄在低镉胁迫下的响应机制,为进一步明确葡萄植株对镉的吸收及其调控机制,提供理论参考。同时为生产中阳光玫瑰葡萄砧木选择、改良葡萄品质提供理论支撑。
1 引言
镉(cadmium, Cd)对于植物来说是非必需的生理元素。与其他重金属相比,Cd是一种相对可移动的元素,极易被植物根系吸收和转运,即使很低浓度的Cd也会对生物体造成危害(Chen and Kao, 1995)。Yang等(2018)调查发现,36.7%的土壤样品中Cd浓度超出了中国土壤环境质量标准(GB15618-1995)的II级值(0.3 mg/kg)。因此,研究植物如何应对镉胁迫是十分必要的。李小红(2018)发现嫁接可以提高矢富罗莎葡萄植株对Cd胁迫的耐受性,但葡萄植株对Cd胁迫的响应机制不仅与Cd处理的浓度和时间相关,还与品种密切相关,而Cd对阳光玫瑰鲜食葡萄的影响,还未见报道,同时,不同砧木嫁接阳光玫瑰葡萄对Cd的富集和抗性特点、嫁接后阳光玫瑰葡萄在Cd胁迫下的生理生化反应以及转录和代谢水平的变化,尚不清楚。
代谢组学的研究对象是生物系统(由细胞、组织或生物构成)受到刺激或干扰后,其代谢物的变化或随时间的变化规律的一门科学(许国旺等,2007)。植物在不利的环境中时,非生物胁迫阻碍了植物生长和发育,所以,植物在目前应激条件下,代谢网络须重新配置,维持所需代谢,以及利用新稳态适应逆境(Kotak et al., 2007)。因此运用代谢组学探讨植物对非生物胁迫的应答机制和代谢网络调控有着重要意义。在高浓度镉胁迫下,Bailey等人(2003)对麦瓶草(Silene cucubalus)细胞进行了代谢组学分析,同时还发现苹果酸,醋酸盐含量明显增加,但谷氨酸及部分支链氨基酸的含量有下降的趋势。Kieffer等(2009)通过蛋白质组和代谢组结合的研究,发现白杨在镉胁迫过程中色素含量,碳水化合物含量均发生变化,光合作用受到不同程度的影响,在植株的生长发育期间,光合作用产物以己糖或者其他复杂糖类的形式储存,从而起到渗透调节的效果。同时,最近发现紫花苜蓿在镉胁迫条件下会提高脯氨酸、游离氨基酸含量,可能是紫花苜蓿抗镉毒害(Yang et al., 2019)的关键环节。
因此,本试验以三种不同砧木(生产常用)嫁接的阳光玫瑰葡萄为试验材料,用不同浓度的Cd处理土壤,研究Cd胁迫对葡萄代谢组的影响;通过比较三种不同砧木对Cd的耐受性,以及Cd处理后葡萄叶片的Cd积累,通过代谢组分析,对三种砧穗组合响应镉胁迫进行差异代谢物分析,主要包括差异代谢物的鉴定、PCA分析、KEGG通路富集分析等,探究三种砧穗组合在代谢水平的差异及葡萄响应镉胁迫的重要代谢通路。上述研究上述研究对于了解Cd胁迫对葡萄基因的表达和代谢的调控机制,具有重要意义,可为生产上通过选择适宜的砧木、减少重金属危害、促进优质葡萄生产,提供理论支撑。
2 材料与处理
2.1 试验材料
本研究中使用的葡萄材料为嫁接一致的一年生阳光玫瑰,所选的砧木品种分别为SO4、5BB和3309C三种常见的阳光玫瑰砧木品种,嫁接苗采用简写形式分别为SM/SO4、SM/5BB和SM/3309C。
葡萄园土过筛,每盆称重20kg,将CdCl2按需换算为Cd浓度梯度。Cd浓度梯度为0 mg/kg、0.2 mg/kg、0.4 mg/kg、0.8 mg/kg、1.6 mg/kg、3.2 mg/kg,后将每盆所需CdCl2溶于500 ml蒸馏水,边铺土边喷含Cd溶液,最后搅拌3-5次,保证充分混匀。每个Cd浓度处理的每种砧木设置3个生物学重复,以0 mg/kg的为对照组,共计54盆。定植28天后,待叶片长到6-8片叶时,取长势好、成熟、完整的葡萄叶片于12 小时收集样品,立即在液氮中冷冻,并储存在-80℃冰箱,用于非靶向代谢组分析。
2.2 代谢组分析
2.2.1 样本制备与上机分析
代谢组样本与上述转录组相同,分为12 组,每组有6 个生物学重复,共计72个样品进行代谢组样本制备,上述过程由上海美吉生物有限公司完成。具体设备、试剂及参数如下表所示:
2.2.2 差异代谢物分析
本试验中差异代谢物的选择是在美吉生物云平台上根据OPLS-DA模型所得变量权重值(VIP)和student's t检验p值来确定,即P<0.05且VIP>1的代谢物为差异代谢物。获得的差异代谢物通过比对HMDB 4.0数据库,进行分类与统计,用Python软件包scipy.stats进行通路富集分析,并通过Fisher精确检验获得与镉胁迫相关的生物学途径。具体分析软件详情如下表所示:
3.结果与分析
3.1三种砧穗组合代谢组学分析
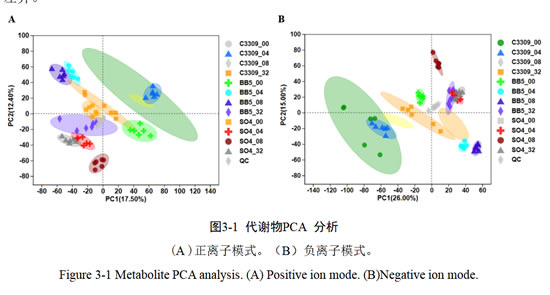
3.1.1 主成成分分析
样本经液相色谱分离条件得到的数据, 采用主成成分分析,观察各样本之间整体分布趋势。如图3-1,选取前两个主成分进行作图。结果表明,QC样品分组良好,表明生物分析和数据质量是良好的,说明组间分离是由于组间的差异变量,而非分析过程中的差异造成的。第一主成分(PC1维图)上正、负离子模式下的贡献率分别为17.50%、26.00%,12组样本之间通过PCA分析能被分离。在第二主成分(PC2维图)上可以看出、每个胁迫中的样本较好的聚在一起、表明试验过程中生物学重复之间的差异较小、组内变异度较小。总体来说,该PCA分析结果能够解释样本在代谢水平的差异。
3.1.2 差异代谢物鉴定
本实验选取72个样品,分为12组进行代谢组研究,每组有6个生物学重复。基于LC-MS检测平台和自建数据库共检测到742个代谢物。其中正离子模式下共检测到426个代谢物、负离子模式下共检测到316个代谢物。以P<0.05、fold change>1 或fold change<1和VIP(Variable Importance InProjection)>1为标准进行差异表达代谢物(Differentially expressed metabolites、DEMs)的筛选。
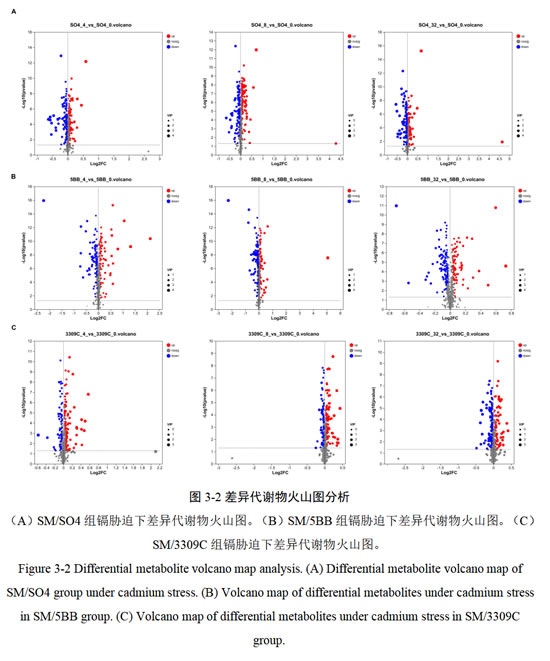
与对照相比,SM/SO4组在0.4mg/kg的镉胁迫下共检测到190种显著差异代谢物,92种代谢物上调,如脂酰GMP(Lipoyl-GMP、1.36倍),98种代谢物下调,如4-硫酸多巴胺(Dopamine 4-sulfate、0.69倍);在0.8mg/kg的镉胁迫下共检测到206种显著差异代谢物,102种代谢物上调,如精氨酸精氨酸(Arginyl-Arginine、19.59倍),104种代谢物下调,如脱氧胞苷(Deoxycytidine、0.64倍);在3.2mg/kg的镉胁迫下共检测到189 种显著差异代谢物,82种代谢物上调,如精氨酸精氨酸(25.09倍),107种代谢物下调,如抗坏血酸(L-Ascorbic acid、0.78倍)(图3-2A)。
与对照相比,SM/5BB组在0.4mg/kg的镉胁迫下共检测到185种显著差异代谢物,86 种代谢物上调,如异牡荆素(Isovitexin、2.49倍),99种代谢物下调,如佩塔辛甙(Petasinoside、0.61倍);在0.8mg/kg的镉胁迫下共检测到191种显著差异代谢物,80 种代谢物上调,如精氨酸精氨酸(33.6倍),111种代谢物下调,如佩塔辛甙(0.60倍),在3.2mg/kg的镉胁迫下共检测到199种显著差异代谢物,88 种代谢物上调,如牡丹素3-(6''-对香豆基葡糖苷)(Peonidin3-(6''-p-coumarylglucoside)、1.66倍),111种代谢物下调,如佩塔辛甙(0.61倍)(图3-2B)。
与对照相比,SM/3309C组在0.4mg/kg的镉胁迫下共检测到201种显著差异代谢物,133 种代谢物上调,如4-硫酸多巴胺(1.41倍),68种代谢物下调,如牡丹素 3-(6''-对香豆基葡糖苷)(0.67倍);在0.8mg/kg的镉胁迫下共检测到159种显著差异代谢物,98 种代谢物上调,如佩塔辛甙(1.30倍),61种代谢物下调,如6'-硫酸软骨素(Chondroitin 6'-sulfate、0.87倍),在3.2mg/kg的镉胁迫下共检测到168种显著差异代谢物,71种代谢物上调,如苯乙酸乙酯(1.21倍),97种代谢物下调,如4-(2-硝基乙基)苯基芋甲苷(4-(2-Nitroethyl)phenyl primeveroside、0.72倍)(图3-2C)。
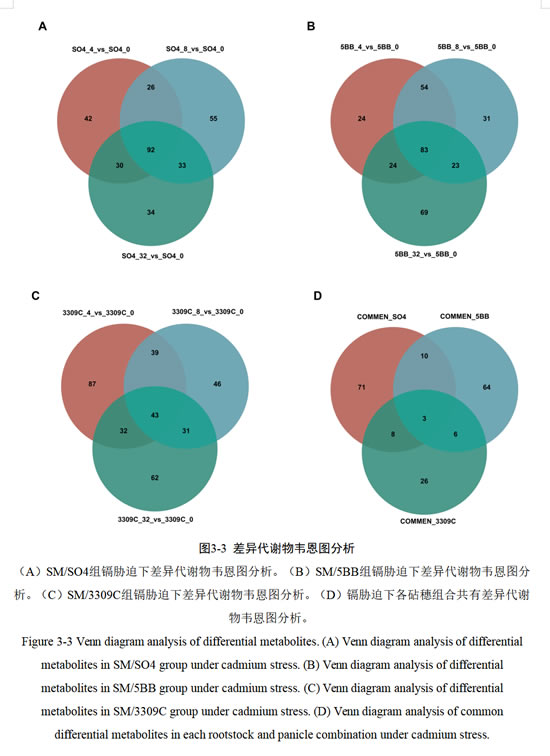
利用韦恩图分析葡萄三种砧穗组合、不同镉浓度胁迫的差异代谢物情况(图3-3),结果表明,SM/SO4组共有的差异代谢物数量为92;SM/5BB共有的差异代谢物数量为83;SM/3309C共有的差异代谢物数量为43。同时对不同砧穗组合的共有差异代谢物进行韦恩图分析,发现SM/SO4组特有的差异代谢物有71个,SM/5BB组特有的差异代谢物有64个,SM/3309C组特有的差异代谢物有26个。
3.4.3 差异代谢物通路富集分析
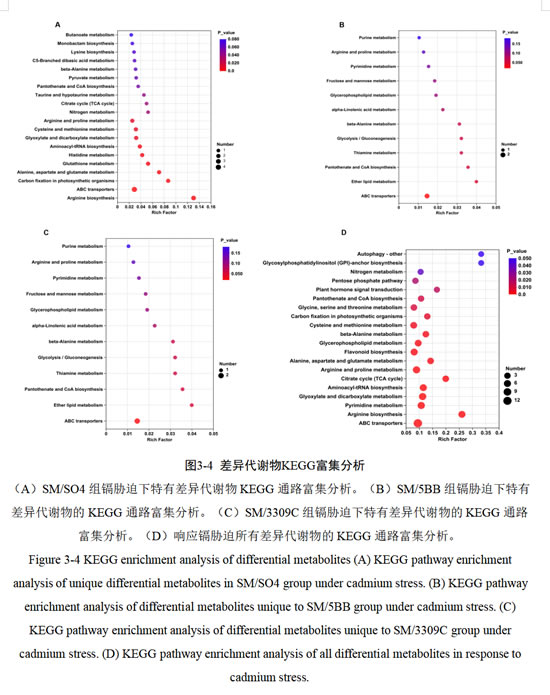
在本研究中,我们利用KEGG 数据库,对上述特有差异代谢物进行代谢通路富集分析,结果显示于气泡图中(图3-4)。可以发现,三组砧穗组合在低镉胁迫条件下的代谢通路变化具有很大差异性。SM/SO4组的特有差异代谢物共涉及27种代谢通路,其中前13条通路被显著富集,依据富集分析的P 值进行排序,显著富集且排名前5的代谢通路分别是精氨酸生物合成,ABC转运蛋白,光合生物的固碳作用,丙氨酸、天冬氨酸和谷氨酸代谢,谷胱甘肽代谢(图3-4A);SM/5BB组的差异代谢物共涉及12种代谢通路,其中前2条通路被显著富集,根据P值进行排序分别是ABC转运蛋白,乙醚脂质代谢(图3-4B);SM/3309C组的差异代谢物共涉及7种代谢通路,其中前5条通路被显著富集,根据P 值进行排序分别是自噬-其他,糖基磷脂酰肌醇(GPI)-锚生物合成,C5支化二元酸代谢,磷酸戊糖途径,甘油磷脂代谢(图3-4C)。同时将响应镉胁迫的所有差异代谢物进行KEGG富集分析,发现显著富集的通路有25条,其中前5条(根据P值排序)为精氨酸生物合成,ABC转运蛋白,乙醛酸和二羧酸代谢,嘧啶代谢,氨酰-tRNA生物合成(图3-4D)。
综上所述,SM/SO4组大量差异代谢物显著富集在氨基酸代谢,ABC转运蛋白,碳水化合物代谢等通路;SM/5BB组差异代谢物显著富集ABC转运蛋白,乙醚脂质代谢通路;SM/3309C组差异代谢物显著富集在磷酸戊糖途径,甘油磷脂代谢,自噬-其他通路,同时响应镉胁迫的所有差异代谢物显著富集在氨基酸代谢,碳水化合物代谢,ABC转运蛋白,核苷酸代谢,脂类代谢,其他次级代谢物的生物合成等通路。可以发现在代谢水平上,SM/SO4和SM/5BB对镉胁迫的响应更为相似,都显著富集在ABC转运蛋白。
4 结论
本研究围绕葡萄在种植中镉污染的基本问题,开展嫁接葡萄响应镉胁迫相关机制研究。利用三种常见砧木嫁接的阳光玫瑰葡萄,结合三种砧穗组合镉处理后叶片组织的代谢组检测结果,探究镉胁迫下接穗叶片的基因代谢谱的变化。主要结果如下:
在代谢水平上,SM/SO4特有差异代谢物显著富集在氨基酸代谢、ABC转运蛋白、碳水化合物代谢通路;SM/5BB特有差异代谢物显著富集在ABC转运蛋白、乙醚脂质代谢等通路;SM/3309C组特有差异代谢物显著富集在磷酸戊糖途径、甘油磷脂代谢、自噬-其他等通路。同时响应镉胁迫的所有差异代谢物显著富集在氨基酸代谢、碳水化合物代谢、ABC转运蛋白、核苷酸代谢、脂类代谢、其他次级代谢物的生物合成等通路。