熟期调控
李胜 曹卓 张德辉 刘建伟 陈勇 陈文婷 杨国顺 白描
摘要:通过二次果进行错季栽培,调控上市时间,是解决目前我国主栽鲜食葡萄品种‘阳光玫瑰’季节性相对过剩的重要手段。错季栽培过程中,关键难点在于二次果成花困难。为解决此问题,本研究采用矮壮素(CCC)+苄氨基嘌呤(6-BA)、矮壮素+胺鲜酯(DA-6)、缩节胺(MC)+苄氨基嘌呤和缩节胺+胺鲜酯等4种处理组合,分别试验了其对主梢和副梢上的冬芽,以及副梢上夏芽的花芽分化的影响以及促花效果。结果发现,相比于对照,各处理都显著提高了冬芽和夏芽的成花率,其中主梢上冬芽的成花率达到90%以上,副梢上冬芽的成花率达到80%以上,不同处理间无显著差异;副梢上夏芽的成花率达到60%以上,以CCC + DA-6处理效果最好,成花率达到94.12%。本研究选择了不同类型的芽萌发后的枝条作为二次果结果枝,应用不同植物生长调节剂组合,探索出了有效的二次果促花方案,对‘阳光玫瑰’葡萄的二次果生产和成熟期调控具有重要的指导意义。
关键词:葡萄;阳光玫瑰;错季栽培;二次果;促花;花芽分化
‘阳光玫瑰’葡萄属欧美杂交种,具有肉质脆硬、玫瑰香味、外观优美、品质上乘、抗病性强、不易裂果、耐运输等优点,深受人们喜爱,目前已经成为我国主栽品种之一。但是由于产量过剩和集中上市的原因,‘阳光玫瑰’葡萄出现了季节性相对过剩的问题,以至于近几年来其价格持续走低,经济效益严重下滑。调控葡萄成熟期可以有效解决季节性相对过剩的问题,而采用收获二次果的错季栽培方式,不仅可将葡萄的成熟期延后,实现葡萄错峰上市,提高经济效益,同时还能在一次果因受自然灾害而欠收的情况下,有效减少经济损失。国内20世纪30年代时就开始尝试葡萄多次结果技术,通过修剪管理和植物生长调节剂处理在一些易成花的葡萄品种中成功获得了二次果。研究发现通过诱发冬芽或夏芽萌发,于当年生长结实,可以在一年内获得一次、二次、三次甚至更多次数的产量收成[7]。然而,‘阳光玫瑰’的二次果成花难的问题还未解决[11],导致其实现二次果错季栽培较为困难。
葡萄的芽为腋生芽,有三种类型:夏芽、冬芽和潜伏芽。夏芽最早从侧生分生组织发育而来,当季可萌发产生副梢;冬芽形成于夏芽副梢先出叶的叶腋中,包含一个主芽和两个或多个较小的副芽,一般需要经历休眠后在第二个生长季才能萌发;保持休眠或隐藏多年的芽为潜伏芽。夏芽花芽分化时间短,通常不结果实,但是在一些成花能力强的品种中也能利用夏芽生产二次果;冬芽生长季节处于休眠状态,冬芽主芽一般将成为下一年产量的主要来源。冬芽中花形成的关键阶段包括第一季的诱导、起始和早期分化以及第二季的芽萌发后分化阶段。葡萄的花芽分化一般指从成花诱导开始到花器官分化的过。Noyce P将葡萄冬芽的花序原基发育分为Stage 0~7。本课题组前期发现,芽中花序原基发育到一定的阶段(通常是Stage 5b之后),就完成了未分化原基向花序原基的分化,此阶段后让芽萌发就能够保证其萌发后枝条上有花。由于葡萄芽具有异质性,不同类型和来源的芽成花能力存在差别。
在葡萄花芽分化过程中,诸多的外部环境(非生物)因素和内部(生物)因素都会对其造成直接或间接的影响,其中内源激素平衡是葡萄成花的重要影响因素。研究表明在葡萄中细胞分裂素(cytokinin,CTK)/赤霉素(gibberellin,GA)比值越大,越有利于成花。而矮壮素、缩节胺、多效唑、烯效唑、调环酸钙等多种矮化剂对赤霉素的合成具有抑制作用。6-BA和DA-6都是细胞分裂素类的植物生长调节剂,其中6-BA可用于促进植物细胞生长分裂、侧芽出现、基枝形成、开花结果;DA-6能够调节植物体内生长素和细胞分裂素水平,能够提高植物光合能力,具有促进细胞分裂和生长,促进开花、座果和加快植物果实成熟的作用。
为解决‘阳光玫瑰’葡萄错季栽培的二次果成花困难的问题,本研究设计出利用不同植物生长调节剂组合促进主梢上的冬芽、副梢上的冬、夏芽结果三种促花方案,探索不同植物生长调节剂组合对不同结果芽的花芽分化的影响及其促花效果。
1 材料与方法
1.1 试验材料
本研究于2023年4月—2023年9月期间开展预试验,于2024年4月—2024年9月开展试验。预试验与试验都在长沙县湖南农业大学干杉试验基地(113°196′E,28°132′N,‘阳光玫瑰’葡萄树龄为8年,飞鸟式架形,株行距1.8 m×2.8 m)中开展,每个处理选择3棵树重复。从萌芽至成花期间以促根肥(海藻肥、腐殖酸和微生物菌剂)和壮条肥(30% N,10% P,10% K)为主。期间在常规病虫害防治的基础上,在冬芽和夏芽萌发后重点注意新梢防虫。
植物生长调节剂及其对照选择:500 mg·L-1 矮壮素(CCC)、25 mg·L-1 苄氨基嘌呤(6-BA)、1000 mg·L-1 缩节胺(MC)、50 mg·L-1 胺鲜酯(DA-6)、清水对照(CK)。
试剂选择:国光牌矮壮素、苄氨基嘌呤、甲哌鎓(缩节胺)、胺鲜酯,四川润尔科技有限公司。
(一)1.2 试验方法
本研究根据湖南地区气候情况判断当地‘阳光玫瑰’最晚成熟期为11月底,故设计利用主梢上的冬芽结二次果将‘阳光玫瑰’成熟期延后至10月底;利用二次夏芽副梢结二次果将‘阳光玫瑰’成熟期延后至10月~11月间;利用副梢上的冬芽结二次果将‘阳光玫瑰’成熟期延后至11月底。又根据‘阳光玫瑰’葡萄的生长周期,推算出冬芽短截时期:6月初短截主梢来促进其上的冬芽萌发,可以将‘阳光玫瑰’葡萄的成熟期推迟到10月底;7月初短截副梢来促进其上的冬芽萌发结果,可以将‘阳光玫瑰’葡萄的成熟期推迟到11月底。
研究中所有处理均用药剂喷雾方式开展,其中处理叶片时用喷雾器喷叶背面,处理芽时从芽上部喷雾,形成一层液膜为宜。
(二)1.2.1利用主梢上的冬芽结二次果
主梢长至5片叶时,去除其上的一次花序。主梢长至10片叶时,留8片叶将其摘心,并将其上的夏芽副梢全部留1片叶绝后摘心。待主梢摘心后,分别用清水、CCC + 6-BA、CCC + DA-6、MC + 6-BA和MC + DA-6处理全株叶片,处理共计3次,每次处理间隔7天。5月31日,取样主梢第四节位冬芽统计其的理论成花率,以判断第三节位冬芽的花芽分化情况。6月1日,于主梢第三节位处短截,并用1.5%单氰胺涂抹该节位冬芽,让其萌发结果。
(三)1.2.2 利用副梢上的冬芽结二次果
主梢长至5片叶时,去除其上的一次花序。待主梢长至7片叶时,分别用清水、CCC + 6-BA、CCC + DA-6、MC + 6-BA和MC + DA-6处理全株叶片1次。待主梢长至8片叶时留3片叶将其短截,并将顶端夏芽副梢留作二次果的结果母枝,将其余夏芽副梢留1片叶绝后摘心。然后自顶端夏芽副梢长至5片叶时开始,分别用上述1.2.1中组合处理全株叶片,处理共计2次,每次处理间隔7d,以促进副梢冬芽的花芽分化。待顶端夏芽副梢长至8片叶时将其留6片叶摘心。6月30日,取样顶端夏芽副梢第三节位冬芽并统计其的理论成花率,判断夏芽副梢第二节位冬芽的花芽分化情况。7月1日于夏芽副梢第二节位处短截,并用1.5%单氰胺涂抹该节位冬芽,让其萌发结果。
(四)1.2.3 利用副梢上的夏芽结二次果
主梢长至5片叶时,去除其上的一次花序,并分别用清水、CCC + 6-BA、CCC + DA-6、MC + 6-BA和MC + DA-6处理全株叶片和其上夏芽,来控制枝梢和夏芽生长。待主梢长至7片叶时留3片叶将其短截,并将顶端的第3节位夏芽副梢保留,将其余副梢留1片叶绝后摘心,分别用上述1.2.1中组合处理全株叶片和保留的顶端夏芽副梢。待顶端夏芽副梢长至4片叶时留2片叶将其摘心,并保留顶端的第二次夏芽,同时再次用上述1.2.1中组合处理全株叶片和顶端的二次夏芽,最后二次夏芽萌发形成的二次夏芽副梢作为结果枝。
(五)1.3 冬芽萌发率统计
利用冬芽结二次果的方案中,短截后用1.5%的单氰胺涂抹短截节位的冬芽,并于30 d后统计该节位冬芽的萌发率。
(六)1.4 成花率统计
(七)1.4.1 理论成花率
利用冬芽结二次果的方案中,短截前1 d在每棵树上随机挑选15个枝条,取短截节位临近上一节位的冬芽,在体式显微镜下剥出其花序原基,按照Noyce P等[16]提供的方法判断花序原基发育所处阶段,统计达到stage 5b后的芽所占的比例作为理论成花率。
理论成花率(%)=达到stage 5b及以后阶段 的芽数目/取样芽总数×100
(八)1.4.2 实测成花率
统计短截节位冬芽萌发后的成花率(利用冬 芽结果的方案),以及二次夏芽副梢的成花率(利用夏芽结果的方案)作为实测成花率。在每棵树上花序大小为5 cm有3个及以上分支时进行实测成花率统计。实测成花率(以每株为单元) 计算公式如下,结果为平均值:
实测成花率(%)=短截萌发后带花序的枝条数/短截后萌发的枝条总数×100
(九)1.5 数据分析
试验数据显著性差异用IBM SPSS Statistics 26软件进行分析;用Microsoft Excel 2016进行数据分析和处理。
2 结果与分析
(十)2.1 理论成花率
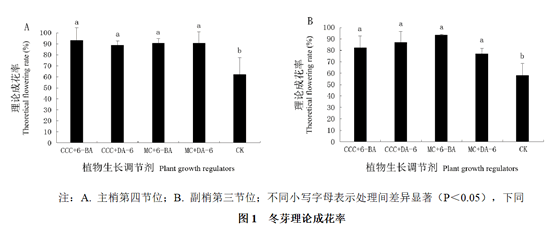
在短截的前一天(5月31日),统计了主梢上第四节位冬芽的理论成花率,结果显示,对照的冬芽理论成花率为62.22%,而CCC + DA-6处理后冬芽的理论成花率为88.89%,其余3种组合处理后冬芽的理论成花率都超过了90%,处理间无显著差异,而各处理与对照均存在显著性差异(图1A)。
6月30日统计了副梢上第三节位冬芽的理论成花率,结果显示,对照的冬芽理论成花率为57.78%,而MC + DA-6处理后成花率接近80%,CCC + 6-BA、CCC + DA-6和MC + 6-BA处理后成花率达到80%以上。副梢第三节位冬芽的理论成花率在各处理间无显著性差异,但均显著高于对照(图1B)。
(十一)2.2 冬芽萌芽率与实测成花率
(十二)2.2.1 冬芽萌芽率
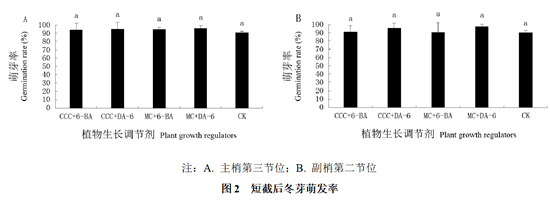
经过统计发现,短截后主梢的第三节位冬芽和副梢的第二节位冬芽的萌发率都达到90%以上,且各处理间及其与对照间都不存在显著性差异(图2)。
(十三)2.2.2 主梢上冬芽的二次成花率
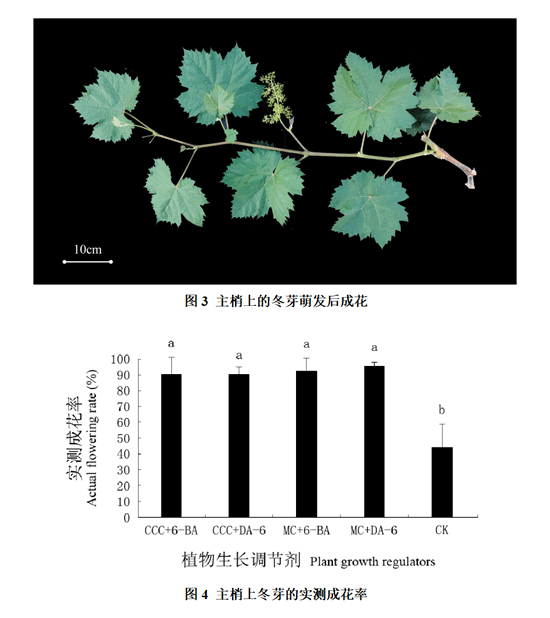
统计主梢上第三节位冬芽的实测成花率发现:对照的冬芽实测成花率低于50%,而各处理的冬芽实测成花率都高于90%,均显著高于对照,而各处理间差异不显著(图3、4)。
(十四)2.2.3 副梢上冬芽的二次成花率
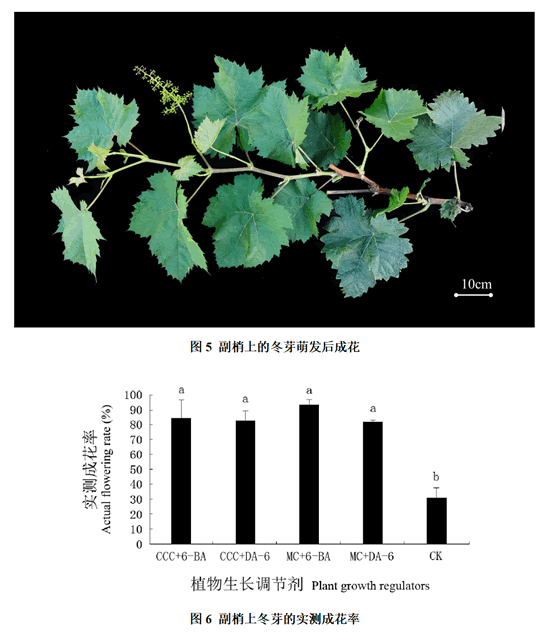
统计副梢上第二节位冬芽的实测成花率发现:对照的冬芽实测成花率仅为30.96%,而各处理的冬芽实测成花率都高于80%,其中MC + 6-BA处理的冬芽实测成花率超过了90%。各处理的冬芽实测成花率均显著高于对照,但各处理间差异不显著(图5、6)。
(十五)2.3 二次夏芽副梢的成花率
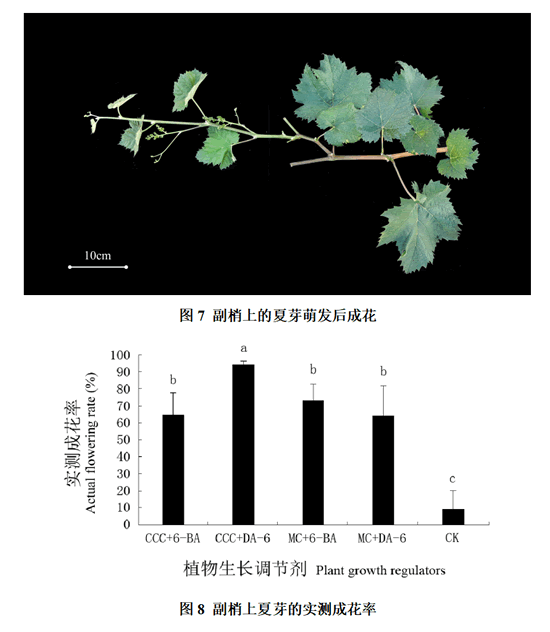
统计二次夏芽副梢的实测成花率发现(图7、8):对照中二次夏芽副梢的实测成花率极低,且重复间的差异很大,成花非常不稳定;CCC + 6-BA、MC + 6-BA和MC + DA-6处理后二次夏芽副梢的实测成花率分别达到64.78%、73.02%和64.19%;而CCC + DA-6处理后二次夏芽副梢的成花率最高,达到94.12%,且该处理的重复间差异较小,成花较稳定。经过分析发现,二次夏芽副梢实测成花率各处理都显著高于对照,CCC + 6-BA、MC + 6-BA和MC + DA-6处理间无显著性差异,CCC + DA-6处理显著高于其他处理。另外,观察发现夏芽上花序相对于冬芽萌发枝条上的花序要小(图5、7)。
3 讨论与结论
根据‘阳光玫瑰’葡萄的生长周期以及课题组前期的错季栽培研究[25-27],可以推算:6月初短截主梢来促进其上的冬芽萌发,可以将‘阳光玫瑰’葡萄的成熟期推迟到10月底;7月初短截副梢来促进其上的冬芽萌发结果,可以将‘阳光玫瑰’葡萄的成熟期推迟到11月底。本研究经过取样上一节位冬芽统计其理论花芽率发现,5月31日时主梢上第四节位的冬芽和6月30日时副梢上第三节位的冬芽中的绝大多数已经达到stage 5b,已完成未分化原基向花序原基的分化。研究发现枝条的芽繁殖力从基部到中部逐渐增加,并向顶端再次降低[13]。由此可以推测出这两个时期,相应的主梢上的第三节位冬芽和副梢上的第二节位冬芽中的绝大多数已经形成花序原基,这时短截能够保障冬芽萌发后有较高的成花率,由此说明6月初于主梢上第三节位处短截和7月初于副梢上第二节位处短截,促进相应节位的冬芽萌发后能够保证其枝条上有满足二次果生产的花。
俊鹏等[28]研究发现,低浓度的单氰胺有助于‘阳光玫瑰’葡萄萌芽。本试验中,冬芽经过1.5%的单氰胺处理后萌发情况较好,萌发率都达到90%以上,说明冬芽萌发情况并不会对其实际成花率试验造成较大影响。
本研究发现经过CCC + 6-BA、MC + 6-BA和MC + DA-6四种植物生长调节剂组合处理后,主梢和副梢上的冬芽以及二次夏芽副梢的成花率都显著高于对照。这表明赤霉素合成抑制剂和细胞分裂素的组合处理能够显著提高‘阳光玫瑰’葡萄二次果的成花率。另外,由于主梢和副梢上冬芽的实测成花率在各处理组合间并不存在显著性差异,表明所选择的处理组合的调节剂类型、浓度和次数都足够调节葡萄内源的激素水平,达到促进成花的目的。研究发现CCC和MC对GAs合成具有显著的抑制作用,且它们在GAs合成途径中的强烈抑制作用均发生在上游[21, 29]。至于二次夏芽副梢,采用CCC + DA-6处理后实测成花率显著高于其他三种植物生长调节剂组合,分析可能与夏芽花芽分化时间短有关,该处理组合对内源激素水平或成花因素影响更显著。
对比植物生长调节剂组合处理后冬芽和夏芽的成花情况,发现冬芽的成花率明显好于夏芽。分析其原因可能与夏芽发育和萌发速度较快有关,这导致了成花孕育时间较短,这也体现在二次夏芽副梢同一处理的重复间实测成花率误差较大上。但是经过植物生长调节剂组合处理后,二次夏芽的成花率显著提高,有力地证明了赤霉素合成抑制剂和细胞分裂素组合能够有效促进‘阳光玫瑰’葡萄的花芽分化。在实际生产上,70%以下的实测成花率难保证葡萄有足够的产量,同时飞鸟式架形的生产园利用夏芽结果时后期会存在枝条生长空间不足的问题。综合来讲,利用‘阳光玫瑰’葡萄夏芽来生产其二次果具有非常大的不确定性,经过比较发现冬芽更加具有生产二次果的潜力。
对比植物生长调节剂组合处理后主梢和副梢上的冬芽成花的效果,发现除MC + 6-BA外其余三个处理后主梢上的冬芽成花率都高于副梢上的冬芽。且利用副梢上的冬芽生产二次果,整个操作流程相比主梢上的冬芽更为复杂。又考虑到冬芽在当季处于休眠,促萌时间可以调节,即可以利用主梢上的冬芽将二次果延后至11月底,其效果完全可以替代副梢上的冬芽。综合来看利用主梢上的冬芽生产二次果的方案明显更佳。而在本研究中利用副梢上的冬芽和二次副梢生产二次果的方案探究,可以更有力地说明赤霉素合成抑制剂和细胞分裂素组合对‘阳光玫瑰’花芽分化的显著促进作用。
本研究在探究‘阳光玫瑰’葡萄二次果促花技术时发现,在萌芽后的花序发育过程中会出现花序分枝少、花序卷曲、“跑花”等问题。这可能与湖南地区二次花发育过程中的高温有关。同时也可能与使用CCC和MC两种植物生长调节剂处理后,尤其连年处理后,树势变差,树体营养缺乏有关。如何缓解促花过程中产生的不良影响,还需要进一步探究。
综上,本研究通过CCC + 6-BA、CCC + DA-6、MC + 6-BA和MC + DA-6四种植物生长调节剂组合处理,显著提高了主梢和副梢上的冬芽以及二次夏芽副梢的成花率,表明赤霉素合成抑制剂和细胞分裂素的组合能够有效促进‘阳光玫瑰’花芽分化。同时经过植物生长调节剂组合处理,主梢上第三节位冬芽的实测成花率超过90%,副梢上第二节位冬芽的实测成花率超过了80%,成功解决了‘阳光玫瑰’葡萄二次果成花率低的问题。本研究探索出的利用主梢上的冬芽结二次果的促花方案,能够有效获得具有生产意义的二次花,非常有希望获得良好品质的二次果,对‘阳光玫瑰’葡萄的二次果生产和成熟期调控具有重要的指导意义。