酿酒葡萄品种改良岗位
摘要:本研究以酿酒葡萄‘赤霞珠’和‘西拉’为亲本,对其正反交 519 株杂交后代展开研究,旨在揭示果实主要性状的遗传变异规律,为酿酒葡萄杂交育种提供理论支撑。研究结果显示,正反交后代果实平均发育期为 96 d,低于亲中值 105 d,介于双亲之间,整体趋中遗传且主要倾向于‘西拉’。果穗重平均值 87.01±66.40 g,低于亲中值,表现出小果穗遗传倾向,主要倾向于‘赤霞珠’;穗形指数在超高亲与超低亲中占比均超 70.00%,后代分离广泛。果粒重均值高于亲中值,呈大果粒遗传趋势,倾向于‘西拉’,果形指数变异小,倾向于‘赤霞珠’。可滴定酸含量正交后代在 5.96—8.67g・L-1、反交后代在 5.13—12.43g・L-1 广泛分离,遗传倾向于低酸亲本‘西拉’ 。研究明确了‘赤霞珠’与‘西拉’正反交后代果实主要性状的遗传倾向,为酿酒葡萄杂交亲本的科学选配和新品种选育提供了重要参考,对推动我国酿酒葡萄产业发展具有重要意义。
酿酒葡萄人工杂交是有目的的控制花粉和选择亲本的品种来获得杂交种子,国内酿酒葡萄育种进展我国早在 20 世纪 50 年代初就开始葡萄杂交育种工作,自 21 世纪以来,育种工作进入了快速发展时期,葡萄品种的选育数量逐年增加。目前我国葡萄酒产业目前正处于迅速发展阶段,截止 2013 年底,葡萄酒生产量已达到世界第六、消费量达到世界第五,但目前我国主栽的酿酒葡萄品种以国外引进为主,如赤霞珠、霞多丽、美乐、西拉、长相思等,缺少具有较高影响力的自育品种。开展具有自主知识产权且适应我国风土的酿酒葡萄新品种选育,对品种创新改良促进我国葡萄产业发展,具有重要的产业意义。酿酒葡萄果实的品质是生产优质葡萄酒的前提,研究酿酒葡萄杂交后代果实品质指标的遗传规律对葡萄果实品质改良、提高育种效率有非常重要的作用。
本研究以‘赤霞珠’与‘西拉’正反交后代为试验材料,进行果实成熟期与主要经济性状遗传倾向分析,阐明其遗传变异规律,一方面可以为酿酒葡萄杂交育种亲本选择提供重要的参考依据,另一方面选育出的优良后代可作为中间或终端优系应用于生产,显著改良品种应用结构。
1 材料与方法
1.1 试验材料
研究材料来源于山西农业大学果树研究所酿酒葡萄育种圃,地理位置为北纬 37 度 23分,东经 112 度 32 分。年平均气温 10.6℃,降水量为 400~600 mm,无霜期 160~180 d,海拔 800 m,年日照时数 2300 h,有效积温3675℃,土壤为砂壤及粉砂壤土,土壤 pH 值 7.8。
研究所用试材为‘赤霞珠’ב西拉’(正交)、‘西拉’ב赤霞珠’(反交)两个正反交组合的 F1 代。通过真假子代的筛选以正交 215 个后代和反交 304 个后代的真子代为试验材料。
1.2 试验方法
1.2.1 指标测定的方法
成熟期测定及方法:在葡萄收获之前,以果实种子颜色的变化与可溶性固形物达到21 °Brix 为成熟期评判标准,并进行采样试验。通过计算开花期到浆果的充分成熟期的发育天数来确定成熟期,进行果实经济性状鉴定。
1.2.2 数据统计分析方法
通过 Excel 对数据进行整理,利用相关计算公式计算变异系数、组合传递力、优势率、广义遗传力等遗传指标,用 Excel 对统计的每个性状的调查数据做正态分布图,观察每个性状的分布规律。
变异系数、组合传递力、优势率等的计算公式如下。
变异系数: CV/%=S/F×100;
组合传递力(遗传传递力): Ta/% = F/P × 100;
优势率:Ha/% = (F–P)/P × 100;
利用方差分析计算广义遗传力:
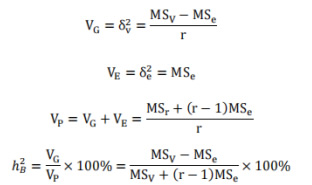
S:标准差;F:后代平均值;P亲本平均值(亲中值);r 随机区组设计中的重复次数;VG:基因型变量;VP:表型变量;VE:环境变量;ℎ��2:广义遗传力。
3 结果与分析
3.1 ‘赤霞珠’和‘西拉’正反交后代成熟期遗传分析
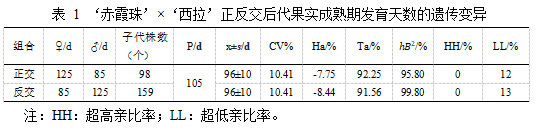
通过 KASP 平台基因分型技术进行真假子代的筛选,得到正交 215 个株系为真子代,占比为 82.06%,反交 304 个株系为真子代,占比为 53.15%。
‘赤霞珠’ב西拉’正反交组合中,组合的广义遗传力分别为 95.80,99.80,表明正反交后代的变异受环境影响小,主要来自遗传效应。如表1所示,亲本‘赤霞珠’果实发育天数为 125 d,亲本‘西拉’成熟期发育天数为 85 d, ‘赤霞珠’ב西拉’的杂交后代果实成熟期调查结果表明,正交后代最晚成熟日期为124 d,正交后代成熟极差为 48 d。后代平均成熟期为 96±10 d,比亲中值早 8~9 d,中晚熟父本遗传优势明显。‘西拉’ב赤霞珠’的杂交后代果实成熟期调查结果表明,反交后代最晚成熟日期为 125 d,反交后代成熟极差为 49 d。反交后代平均成熟期为 96±10 d,早于亲中值 105 d,中晚熟母本遗传优势明显。正反交组合的变异系数均为 10.41%,表明在杂交后代个体间,成熟期表现差异较大。后代平均值均小于亲中值,遗传倾向于中晚熟品种‘西拉’,正反交组合超低亲率为 12.00%和 13.00%,且后代超高亲率均为 0,正反交后代整体呈现趋中遗传趋势,且组合传递力较高,组合在成熟期遗传能力强。
2.2 ‘赤霞珠’和‘西拉’正反交后代主要经济性状遗传分析
2.2.1 ‘赤霞珠’和‘西拉’正反交后代果穗重遗传分析
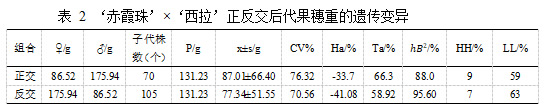
亲本‘赤霞珠’平均穗重 86.52 g,‘西拉’平均穗重 175.94 g。在‘赤霞珠’ב西拉’的杂交后代中,正交后代平均穗重 87.01±66.40 g 小于亲中值 131.23 g,遗传倾向于穗重偏小的亲本,母本遗传优势明显。正交后代中出现超亲现象,超高亲率为 9.00%,超低亲率为 59.00%,变异系数为 76.32%,说明后代分离广泛。正交后代果穗重的广义遗传力为 88.00%,表明果穗重的变异受环境影响小,主要来自遗传效应。在‘西拉’ב赤霞珠’的反交后代中,平均穗重小于亲中值,果穗重量出现小果穗遗传倾向。超高亲率为 7.00%,超低亲率为 63.00%,变异系数为 70.56%,后代同样广泛分离。反交后代果穗重的广义遗传力为 95.60%,变异主要来自遗传效应。
3.2.2 ‘赤霞珠’和‘西拉’正反交后代穗形指数遗传分析
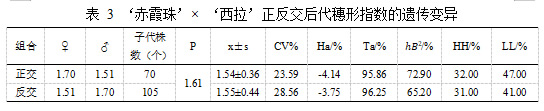
亲本‘赤霞珠’平均穗形指数为 1.70,‘西拉’平均穗形指数为 1.51。在‘赤霞珠’ב西拉’的杂交后代中,正交后代的穗形指数均值小于亲中值,穗形表现为小果穗遗传倾向,从圆锥向短圆锥方向遗传,父本‘西拉’遗传优势明显。超低亲率 47.00%,超高亲率为 32.00%,变异系数为 23.59%,组合传递力高达 95.86%,后代出现超亲现象。‘西拉’ב赤霞珠’反交后代的穗形指数平均值 1.55±0.44 低于亲中值 1.61,有41%后代低于亲本‘西拉’穗形指数,遗传倾向于穗形指数较小的母本‘西拉’。变异系数为 28.56%,组合传递力高达 96.25%。后代短圆锥穗形多可能是受环境影响和亲本非加性效应影响。
3.2.3 ‘赤霞珠’和‘西拉’正反交后代果粒重遗传分析
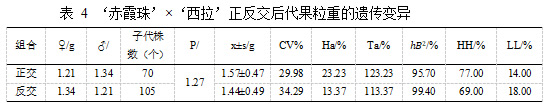
亲本‘赤霞珠’平均果粒重 1.21 g,‘西拉’平均果粒重 1.34 g。在‘赤霞珠’ב西拉’的杂交后代中,正交后代果粒重平均值 1.57±0.47 g 小于亲中值,组合传递力为123.23%,变异系数为 29.98%,超高亲率为 77.00%,超低亲率为 14.00%,具有超亲遗传倾向(见表 4)。在‘西拉’ב赤霞珠’的杂交后代中,反交后代的平均单粒重 1.44±0.49 g 大于亲中值 1.27 g,且大于高亲值,组合传递力达到 113.37%,变异系数为 34.29%,超高亲率为 69.00%,超低亲率为 18.00%,超亲遗传明显,有一定的杂交优势。
2.2.4 ‘赤霞珠’和‘西拉’正反交后代果形指数遗传分析
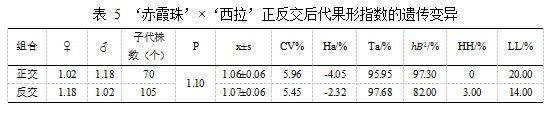
亲本‘赤霞珠’果形指数均值为 1.02,‘西拉’果形指数均值为 1.18。表 5 结果表明,正交后代的果形指数平均值 1.06±0.06 小于亲中值 1.10,与母本果形指数 1.02 相近,表现出较高的倾亲遗传,组合传递力为 95.95%,变异系数为 5.96%,超高亲率为 0,超低亲率为 20%,正交后代果形指数表现出衰退变异,果形有向扁圆遗传趋势。在‘西拉’ב赤霞珠’的杂交后代中,后代的果形指数 1.07±0.06 平均值小于亲中值 1.10,组合传递力为 97.68%,变异系数为5.45%,超高亲率为 3.00%,超低亲率 14.00%。
2.2.5 ‘赤霞珠’和‘西拉’正反交后代可滴定酸含量的遗传分析
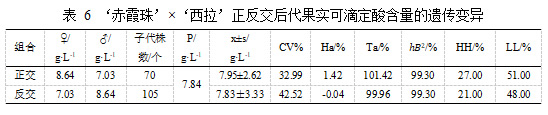
在‘赤霞珠’ב西拉’的杂交后代中,‘赤霞珠’平均可滴定酸含量 8.64 g·L-1,‘西拉’平均可滴定酸含量 7.03 g·L-1。表 6 结果表明,正交后代的可滴定酸含量平均值7.95±2.62 g·L-1 略高于亲中值 7.84 g·L-1,组合传递力为 101.42%,变异系数为 32.99%,超高亲率为 27.00%,超低亲率为 51.00%,有趋向于低酸亲本的趋势,父本遗传优势明显。在‘西拉’ב赤霞珠’的杂交后代中,后代可滴定酸含量平均值接近亲中值,组合传递力达到 99.96%,变异系数为 42.52%,超高亲率为 21.00%,超低亲率为48.00%,有趋于低酸亲本的趋势。
3 讨论
果树性状可分为主基因控制的质量性状与多基因控制的数量性状,为了获得性状的重新组合,前人做过很多不同类型亲本杂交试验,父本和母本通过各自优良特性进行组合,杂交后代出现双亲优良性状的组合,同时也会出现很多超亲现象的结果,甚至会出现双亲的劣势性状,育种过程是选择符合育种目标的个体进行进一步育种,获得性状优良、稳定的新品种。
果实发育期是受多基因控制的数量遗传。正反交组合果实成熟期遗传传递力差异较小,杂种后代遗传变异大,整体呈趋中遗传。在成熟期,后代的遗传与父母本的平均值有密切的关系,表现出超亲遗传。果穗重是葡萄的一个极其重要的经济性状,正反交后代穗形指数的频数分布大致呈正态分布,果穗有普遍偏小的趋势,反交呈正态分布趋势与刘崇怀等的研究反交后代呈正偏分布不一致,酿酒葡萄正反交中超低亲较高,小穗多可能和葡萄的遗传有关,后代果穗变小是个普遍趋势。正反交后代穗形指数是受多基因控制的数量性状。
在葡萄杂交后代果粒重的遗传中,果粒重的遗传符合数量性状遗传。后代的平均值均高于亲中值,杂交后代果粒重呈大果粒倾向遗传,变异的总趋势为超高亲率较多,占 60%以上,这与果粒重量在分级标准及品种的次数分布基本呈正态偏小的分布和郭修武等人研究中杂交后代平均值小于亲中值有差异,整体表现为进化趋势。可滴定酸含量遗传表现为数量性状特点,子代平均值与亲中值相差不大,反交中平均值与亲中值相等。杂交后代中,超低亲率比例较高,后代可滴定酸含量遗传应受非加性效应基因控制,表现下降趋势,低可滴定酸的株系选择潜力较大。正反交后代果实的平均可滴定酸含量接近亲中值。在反交后代呈连续性正偏分布,说明该性状由一对主效基因和多基因共同控制,以加性效应为主,进一步验证了葡萄果实可滴定酸质量分数是由一对主效基因和其他多基因控制的性状的理论。
4 结论
本研究围绕‘赤霞珠’与‘西拉’正反交后代果实主要性状展开,分析其遗传规律,为酿酒葡萄育种提供了关键参考。研究发现,正反交后代果实发育期平均 96d,趋中遗传且倾向于‘西拉’;果穗重性状退化,呈小果穗遗传趋势,倾向于‘赤霞珠’,穗形指数则倾向于‘西拉’ ;果粒重呈大果粒遗传趋势,倾向于‘西拉’,果形指数变异小,倾向于‘赤霞珠’;可滴定酸含量均倾向于低酸亲本‘西拉’。研究结果为酿酒葡萄杂交育种亲本选择提供了重要依据,有助于选育出更优良的品种,推动我国酿酒葡萄产业的发展,解决当前主栽品种依赖国外引进的问题,提升产业竞争力。