酿酒微生物岗位
秦义 陆瑶 杨慧敏 刘延琳
自 20 世纪 80 年代,商业葡萄酒的乙醇含量逐年升高,酒度平均每十年增长约 1%(v/v),上升了3-4% v/v(Dequin et al. 2017; Rolle et al. 2018)。酒度不断升高的原因有两个,一方面,由于全球气候变暖影响了葡萄浆果成熟的不同生理过程。糖分的积累、酚类及芳香类物质的成熟加快,有机酸含量下降,打破了葡萄浆果糖浓度与其他化合物成熟度之间的平衡,使得葡萄酒潜在酒度不断增加。另一方面,消费者对美味、结构饱满、酒体饱满的葡萄酒的需求,推动了市场转向需要采摘非常成熟的葡萄的酿酒风格。因此,现今优质葡萄的含糖量往往偏高,致使其葡萄酒乙醇含量偏高(Mira et al. 2010)。
葡萄酒中乙醇含量过高已成为该行业的棘手问题。葡萄酒的平衡是甜味、酸度、苦味和香气化合物之间的动态平衡。乙醇是香气化合物的载体,过高则会增加苦味和掩盖水果香,并增加甜味、涩味和辣味,从而使感官失衡(Drosou et al. 2022;Englezos et al. 2018; Pham et al. 2020)。从发酵过程角度来看,原料中过高的糖含量可能会影响酵母菌的生长,使发酵缓慢或停滞,过高的酒度又可能使乳酸菌无法生长,使其进行苹乳发酵,从而带来严重的经济损失(Gil et al. 2013, Javier et al. 2022)。除此之外,高乙醇增加了饮酒者的肝脏、消化系统的负担,一定程度上为身体健康带来了风险。另外,高水平乙醇含量的葡萄酒在进出口可能会带来更高的税费支出,使得成本上升,这些因素可能会阻碍葡萄酒的消费,生产酒度适中的葡萄酒是葡萄酒行业亟待解决的问题。
生产低醇葡萄酒的方法主要包括栽培技术、物理技术和微生物技术三个方面。由于栽培技术和物理技术存在成本较高,工艺操作复杂,风味物质损失严重和产生不良副产物等问题,而通过生物技术降低乙醇是较为简单和经济的办法。
1 降低乙醇的方法
生产低醇葡萄酒的方法主要包括主要包括 3 方面,即栽培技术,如葡萄园选址、土壤组成、灌溉策略、砧木、葡萄品种、葡萄产量和叶面积管理等(Burcu et al. 2014);物理技术,减少葡萄或葡萄汁中的可发酵糖,如:葡萄糖氧化酶(Tilloy et al. 2014)、稀释葡萄汁(Cinai et al. 2016)、添加抑制剂(崔艳等 2015)等;从原酒中去除酒精,如:蒸馏、萃取、反渗透等(Dequin et al. 2017);微生物技术,如低产乙醇酵母菌株的选育、酵母混菌发酵、代谢工程改造等。
1.1 栽培方式
一方面,通过减少叶面积可以降低葡萄酒乙醇含量。浆果的糖积累主要是由叶面积与果实重量(LA / FW)之比确定,保证较高的葡萄产量和较低的 LA / FW 比率可以减少糖的积累。因此,座果后减少叶面积能在葡萄酒中降低酒精浓度,也可以通过摘心来实现(Coulter 2013)。在枝条绑缚区或整个葡萄叶幕应用生长调节剂也可以减少葡萄的糖积累(Goold et al. 2016)。另一方面是培育新品种。Aguera(et al. 2010)利用美洲种 Muscadinia rotundifolia 与 4 种不同的欧亚种葡萄 Vitis vinifera 进行杂交,得到的新品种在充分成熟时可以保持较低的含糖量水平。这些新葡萄品种还能抗白粉病和霜霉病。
1.2 物理方法
物理方法减少乙醇含量是目前应用比较广泛的手段,主要是通过葡萄糖氧化酶、替换葡萄汁、添加抑制剂、蒸馏、过滤、渗析以及超临界 CO2萃取等方法减少葡萄醪中的糖、减少乙醇产生以及移除葡萄酒中的酒精(Ciani et al. 2016)。
(1)通过添加葡萄糖氧化酶(GOX),可以在有氧环境中将葡萄糖转变为葡萄糖酸,从而显著降低乙醇的产量(Coulter 2013; Tilloy et al. 2014)。
(2)替换葡萄汁策略提前采摘葡萄进行发酵,可以减少可发酵糖。然而未成熟的果实含糖量低,酿葡萄酒酒度低,会带来酸度高、香气差等品质问题(Rolle et al. 2017)。。
(3)通过一些外源添加的物理手段影响酵母代谢,使其减少或停止乙醇的生产。提前中止发酵,添加 SO2、降温、除去酵母等方法常用于中止发酵,从而降低酒度(李艳等2007)。
(4)移除葡萄酒中酒精,Roole et al.(2018)利用经反渗透处理的葡萄汁酿造葡萄酒,其酒精度降低 1~2%(v/v)。
1.3 微生物方法
考虑到参与葡萄酒酿造过程中酵母种类的多样性以及过去几十年葡萄酒中酒精水平上升的问题。研究者从不同的角度探讨了这个问题:探索非常规酵母的应用(Kelanne et al. 2022),主要集中在非酿酒酵母菌;遗传改良:通过基因工程、随机突变或实验进化,或改变环境酿造条件(新陈代谢抑制剂、好氧)。有氧条件与酵母遗传改良(酵母菌和非酵母菌菌株)相结合是目前最有希望的选择之一。
为了降低乙醇产量,通过代谢工程改造将碳流量转向其他途径以及其他次级代谢产物的产生是较为常见的方法,但代谢改造的同时必须保持氧化还原平衡,避免产生影响葡萄酒感官品质的化合物,并保持酵母的发酵性能,要同时实现这些目标比较困难(Dequin et al. 2017)。
1.3.1 乙醇代谢途径改造
在酒精发酵过程中,从丙酮酸到乙醇有两个酶促反应(图 1-1)。第一个步骤是从丙酮酸到乙醛,由丙酮酸脱羧酶(PDC)催化,在缺乏 PDC 活性的情况下,Crabtree效应被取消,然而这种活性对细胞中乙酰 CoA 的合成和 NAD+的再生是必需的,因此它在酿酒酵母的糖酵解途径中是是必不可少的(Flikweert et al. 1996)。有研究表明,酿酒酵母中编码丙酮酸脱羧酶的基因 PDC1 缺失导致 PDC 活性降低 4 倍,其丙酮酸生成量增加,但乙醇产量并没有降低(Remize et al. 2000)。而敲除参与 PDC 基因表达的转录因子 PDC2 时 PDC 活性为原来的 19%,乙醇含量明显下降,同时甘油和丙酮酸释放增加(Nevoigt et al. 1996)。Cuello et al. (2017)用截短的 PDC2 等位基因替换了不同二倍体酿酒酵母菌中的一个 PDC 等位基因,重组菌的乙醇产量降低了 2% ABV,但乙酸产量升高。
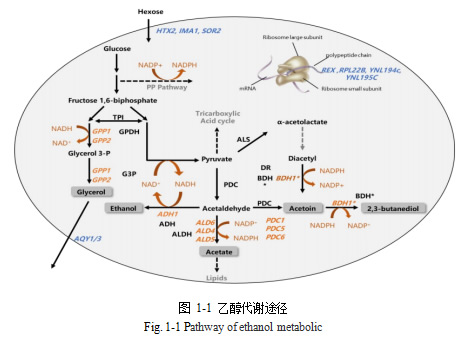
乙醇脱氢酶(ADH)催化酒精发酵过程中乙醇产生的最后一步。研究发现,敲除编码乙醇脱氢酶的基因 ADH1,可减少乙醇生产并增加甘油产量,但重组菌无法在厌氧环境下生长,这种基因改造还导致乙醛和乙酸含量升高,其含量远超商业葡萄酒标准含量(Drewke et al. 1990; Singh et al. 2021)。除了通过沉默或上调内源基因的表达来调节基因表达外,还可以通过外源表达来自其他生物的基因。Dequin(et al. 1994)在酿酒酵母中表达来自干酪乳杆菌的乳酸脱氢酶编码基因 LDH,将糖酵解中形成的丙酮酸转化为乳酸,重组酵母菌同时进行酒精发酵和乳酸发酵,减少 1%的乙醇会增加 15g/L的乳酸。因为两种途径消耗等量 NADH,乳酸形成被等分子量的乙醇消耗所补偿,所以该过程中氧化还原平衡不受影响(Li et al. 2022)。乳酸产量增加会导致总酸升高,可用于低酸原料的酸补偿,但可能会使葡萄酒的不平衡(Vicente et al. 2022)。Yu(et al. 2018)改造了酿酒酵母的代谢以过量产生游离脂肪酸,并进一步证明,通过这种基因编辑,可以用脂肪生成代替酒精发酵作为代谢葡萄糖的主要途径。通过用一个嵌合基因取代所有活跃的己糖转运基因,构建了一株 Crabtree 阴性酿酒酵母,只要进行有氧呼吸该菌株就不会产生乙醇,但其生长极其缓慢(Otterstedt et al. 2004)。
1.3.2 甘油代谢途径改造
甘油对葡萄酒的粘度、酒体和甜度有积极的贡献(Gawel et al. 2007; Paup et al. 2023)。在酵母基因改造策略中,改造甘油合成途径已成为降低乙醇产量的首选。过量表达编码甘油-3-磷酸脱氢酶的基因 GPD1 会显著增加甘油产量(增加 28 g/L)和乙醇的减少(减少 15 g/L),同时会带来过量的乙酸盐和乙偶姻,以达到发酵过程中细胞内的氧化还原平衡(Michnick et al. 1997)。敲除编码乙醛脱氢酶的基因 ALD6 会使乙偶姻产量增加,其浓度过高时呈现出黄油味,给葡萄酒带来负面影响(Remize et al. 2000; Zheng et al. 2020)。基于此,Ehsani(et al. 2009)通过过量表达编码丁二醇脱氢酶的基因 BDH1 来解决乙偶姻产量过高的问题,使乙偶姻在丁二醇脱氢酶的催化下转化为没有明显感官特征的 2, 3-丁二醇,使用重组菌发酵的葡萄酒乙醇产量显著下降,甘油产量超过 26g/L。在缺失基因 PDC 的酿酒酵母中过量表达 GPD1,不仅增强了甘油-3-磷酸的活性,而且消除了在原始菌种直接过量表达 GPD1 所带来的乙酸和乙醛的增加(Nevoigt et al. 1996; Huo et al. 2022)。
由TPI1 编码的丙糖磷酸异构酶时催化1, 6-二磷酸果糖分解产生的两种产物之间相互转化的酶(图 1)。该通路的结构表明 TPI1 缺失会产生等摩尔水平的甘油和乙醇前体物质,但在实际操作中由于氧化还原失衡,重组菌无法在葡萄糖培养基中生长(Compagno et al. 2001;Cordier et al. 2007)。工业上为了提高甘油产量,在 TPI1 缺失菌株中敲除 ADH1,同时过量表达 GPD1 及编码甘油转运蛋白的基因 FPS1 和编码醛脱氢酶的基因 ALD3,获得的重组菌的乙醇产量约为亲本的 25%,甘油含量为亲本的30 倍,但其生长速度大大降低了(Rossouw et al. 2013)。其所需的不同遗产修饰的步骤繁多,因此应用此方面以达到降低乙醇产量存在一定难度。
1.3.3 氧化还原途径
改造乙醇、甘油代谢途径时涉及氧化还原平衡和相关辅因子的补充。因此,一些研究者试图直接作用于氧化还原系统,已达到降低酵母的乙醇产量。通过克隆一个用于再生还原力的 NADH 氧化酶并控制通氧量,获得的葡萄酒乙醇产量显著下降,乙醛、乙酸和乙炔的含量显著上升(Heux et al. 2006)。
1.3.4 其他途径改造
葡萄糖氧化酶处理葡萄汁是降低葡萄酒酒精含量的另一种方法。基于这种方法,Malherbe(et al. 2003)酿酒酵母中外源表达黑曲霉菌中的一个葡萄糖氧化酶基因,用重组菌发酵的葡萄酒乙醇含量降低了 2%,并且重组菌对乳酸菌具有抑制作用。从随机定向诱变产生的 SPT15(编码转录因子)突变体文库中选出一株乙醇产量降低的菌株。这种突变的多效性下调己糖转运和乙醇生产,并上调甘油生产,乙酸没有增加,但突变株生长速度下降(Du et al. 2020)。