石家庄综合试验站
葡萄是我国栽培面积最广的林果树种之一。近年来由于其经济效益明显,葡萄产业已成为许多地区促进经济发展、带动农民增收的重要途径。浆果颜色是葡萄重要的外在品质之一,直接影响果实的经济价值。葡萄颜色复杂多样,从白色到紫黑存在众多着色类型。另外,河北地区夏季雨热同季,连绵阴雨与套袋栽培导致葡萄果实着色不佳。近些年设施栽培发展迅速,成为推动中国葡萄产业发展的新动力。但是设施栽培常常因光照弱造成果实花青素积累不足,着色不良,严重影响葡萄的的经济效益。葡萄果实颜色多样,花青素的积累存在复杂的调控机制,通过研究花青素的积累模式及关键基因的调控途径,促进对花青素的进一步认知,为培育适应的新品种奠定基础。
1 材料和方法
1.1 材料及其处理
试验于2023年6月在河北省农林科学院石家庄果树研究所葡萄示范园塑料大棚内进行。试验树为5年生的摩尔多瓦葡萄,株行距1m*2.2m,厂字架,V型叶幕。果园土壤均为砂壤土,试验期间果园日常管理(挂果量、肥水)保持一致。。
1.2 样品采集
葡萄果实于转色前(青果)、转色初、转色末采集。每处理每次采样100粒果实,注意兼顾果穗的上、中、下各部位,随机选取30粒作为一个重复,设3次重复。剥取果皮,检测花色苷含量。余下锡箔纸包裹,液氮速冻后-80℃保存。
1.3 RNA提取与质量检测
采用天根磁珠法植物总RNA提取试剂盒提取各样品的RNA。1%琼脂糖凝胶电泳与Nanodrop 2000分光光度计检测RNA的浓度及纯度,Agilent Bioanalyzer 2100检测RNA的完整性。
1.4 转录组测序
将提取的样本总RNA送至上海派森诺生物科技有限公司测序并进行文库构建。采用Illumina Novaseq 6000平台对质量合格的文库进行测序。测序得到的图像文件经平台自带软件进行转化,生成FASTQ的原始数据(Raw Data)。去除原始数据中带有接头序列的与低质量的Reads,得到Clean Reads。下载葡萄参考基因组的序列文件和注释文件(http://plants.ensembl.org/Vitis_vinifera),使用HISAT2软件将Clean Reads与葡萄参考基因组进行比对分析。
1.5 差异基因的筛选与功能注释
利用HTseq进行表达水平评估,并采用FPKM值对各处理表达值进行标准化。使用DESeq软件(1.20.0)进行样品组间的差异表达分析。筛选差异表达基因条件为:|log2FC| > 1且P-value<0.05。使用topGO对差异表达基因进行GO富集分析并计算P-value,显著富集标准为P-value < 0.05。使用cluster Profiler(3.4.4)软件进行KEGG 通路富集分析,显著富集标准为P-value < 0.05。
2 结果与分析
2.1 葡萄果实的表型特征变化
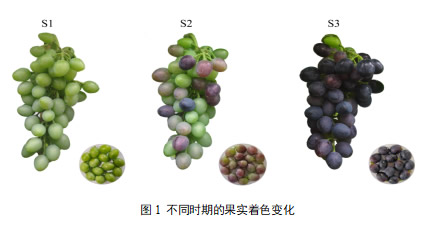
葡萄皮在不同发育阶段的表型差异。在葡萄发育过程中,分别选择了55d(S1)、65d(S2)、78d(S3),建立了葡萄皮中花青素总变化的系统图谱。如图1所示,随着时间的推移,葡萄的颜色逐渐从绿色变成红色、紫黑色。对三个阶段总花青素含量的定量分析也证实了葡萄皮中花青素在整个发育阶段不断积累。S1样品的总花青素含量最低,花青素含量为31.46 ug/g,然后逐渐上升到10345.66 ug/g(S3)。
2.2 果皮在发育过程中的转录组学分析
为进一步阐明葡萄皮花青素合成的分子机制,对3个发育阶段(每个阶段3个生物重复)进行了测序,构建了9个cDNA文库。在去除适配器和低质量的reads后,超过89%的Clean Reads与葡萄参考基因组相匹配(PN40024.v4.dna.toplevel.fa.gz ),获得了约60.75g(Gb)的质控数据,每个样本的平均数据约为6.75 Gb。9个文库的GC含量为45.32%~47.07%,Q30的最低含量为92.75%。上图率为93.28%~94.48%,唯一上图率为89.82 %-91.03%。这些结果表明,测序数据的准确性和质量高度满足了进一步分析的需要。
2.3 差异表达基因的分析
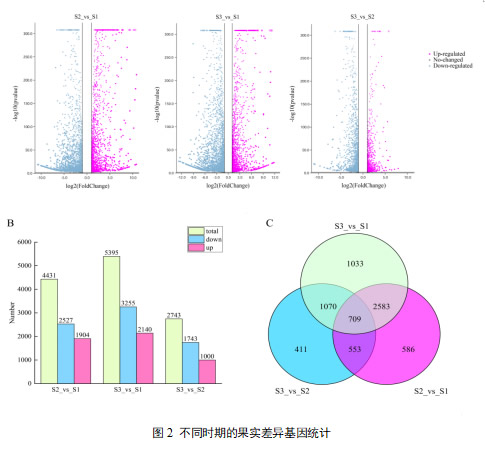
基于|log2Fold Change|≥1与FDR<0.05,共检测到6945个差异表达基因,其中,S2/S1组间有1904个上调和2527个下调;S3/S1组间有3255个下调和2140个上调;S3/S2组间有1000个上调,1743个下调。维恩图显示,在S2/S1、S3/S1和S3/S2阶段之间分别有586、1033和411个独特的DEGs,三组之间共有709个DEGs。DEGs(S3/S1、S2/S1)的数量随着葡萄皮颜色的增加而增加,说明后期参与染色过程的基因较多,而S3/S2 DEGs的数量减少,提示存在持续表达或下调的基因。
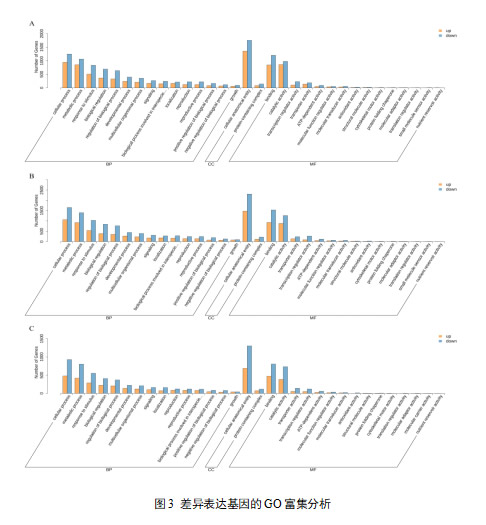
为了更好地了解DEGs在葡萄皮过程中的生物学功能,我们分别采用了GO、KEGG分析。如图5所示,根据GO数据库,DEGs主要富集于41个GO项中,包括22个生物过程,2个细胞组分和16个分子功能。在生物过程类别中,大多数deg被标注为“代谢过程(GO:0008152)”、“细胞过程(GO:0009987)”、“刺激反应(GO:0050896)”和“生物调节(GO:0065007)”、“生物过程调节(GO:0050789)”。在细胞成分类别方面,DEGs仅在“细胞解剖实体(GO:0110165)”和“含蛋白复合物(GO:0032991)”中富集,主要集中在前者。在分子功能类别中,DEGs主要富集于“催化活性(GO:0003824)”、“结合(GO:0005488)”,其次是“转运体活性(GO:0005215)”、“转录调节活性(GO:0140110)”和“atp依赖活性(GO:0140657)”。 此外,为了探索参与花青素生物合成调控的关键基因,我们还对DEGs进行了KEGG注释。KEGG注释显示,S2_vs_S1组间富集133条通路,共富集1410个DEGs;S3_vs_S1组中有1617条在137条通路中富集(补充表S11);S3_vs_S2组中127条通路中有845条富集。根据各组DEGs显著富集的途径统计,四种最显著的途径包括代谢途径(ko01100)、次生代谢物生物合成(ko01110)、植物激素信号转导(ko04075)、植物−病原体相互作用(ko04626)。
2.4 花青素生物合成途径的关键DEGs
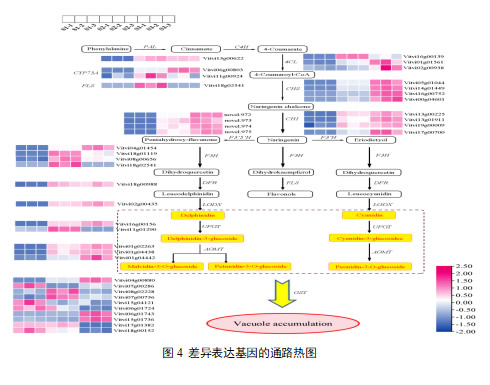
为了探讨三个果皮间花青素生物合成的差异,我们鉴定了花青素合成途径中的DEGs,包括苯丙素生物合成、类黄酮生物合成和花青素生物合成途径。结果显示,有14个结构基因(40个转录本)参与了花青素的生物合成。 在花青素生物合成的早期阶段,共筛选出1个PAL基因、3个4个CL基因、3个CYP73A基因、4个CHS基因和3个CHI基因。PAL(Vitvi13g00622)在S1期低表达,在S2和S3状态下持续高表达。CHS(Vitvi05g01044)在S1期表达量较低,在S2期和S3期表达量迅速增加,成为表达量最高的基因。 柚皮苷转化为二氢山奈酚的过程是F3H基因催化花青素生物合成的关键。1个F3H基因(Vitvi04g01454)在S1期低表达,且随着果实发育而逐渐升高,其他F3H基因(Vitvi18g01119、Vitvi08g00656、Vitvi18g02541)在S1期表达量最低,从S2期到S3期先增加后下降。
F3‘H和F3’5‘H决定了花青素前体成分。S3期F3‘H(Vitvi17g00700)的表达量接近S2期的2倍,S1期的3倍。4个f3’5’H基因在S1状态下几乎不表达,且随着果实的发育,其表达水平逐渐增加。花青素生物合成的后期阶段对花青素的积累也很重要。DFR和ANS在S1期表达量较低,并逐渐增加。1个UFGT(Vitvi16g00156)基因表达上调,另一个UFGT(Vitvi11g01290)基因在S1~S3期先升高后下降。筛选出3个AOMT基因(Vitvi01g02263、Vitvi01g04438、Vitvi01g04442),其表达量从S1逐渐增加到S3。10个GST(Vitvi04g00880, Vitvi07g00286, Vitvi08g02228, Vitvi07g00736, Vitvi15g04121, Vitvi06g01724, Vitvi06g01743, Vitvi15g01736, Vitvi17g01382, Vitvi18g00152)基因具有不同的表达模式。3个GST基因(Vitvi04g00880、Vitvi06g01743、Vitvi15g01736)从S1期到S3期持续增加。
2.5 相关转录因子分析
转录因子是转录调控的核心功能蛋白,在植物的发育过程中起主要的调控作用,在果实花色苷的生物合成也充当重要角色。在不同阶段下,葡萄果皮均有众多转录因子基因差异表达,主要包含MYB、bHLH、ERF、C2H2、NAC、MIKC-MADS、Dof等,表达模式上调下调均有。
3 结论
葡萄果实着色过程伴随着花色苷的积累,转录组数据分析显示不同着色阶段存在众多的差异表达基因,其中大部分花色苷通路的结构基因PAL、CHS、CHI、F3’H、F3’5’H、DFR、UFGT等基因均呈上调表达,尤其是CHS上调表达极高。另外,转录因子在葡萄果实转色过程中发挥重要功能,本研究也发掘众多参与花色苷积累的潜在因子,需要后期开展更多研究进行验证。