鲜食葡萄栽培岗
刘静 陶建敏
摘要:为研究外源赤霉素(GA3)处理对葡萄果形变化的影响,以指形葡萄品种‘金手指’为试验材料,于盛花后3 d使用25 mg /L的GA3溶液对葡萄花序进行处理,清水处理作为对照。观察葡萄果实形态变化,测量果实纵横径并计算果形指数;制作石蜡切片观察果实内部细胞形态差异,测量细胞纵横径并计算细胞指数;利用ESI-HPLC-MS/MS测定48 h和4 d后果实内源赤霉素、生长素和IAA-天冬氨酸(IAA-ASP)含量变化;通过qRT-PCR检测果实中生长素合成和信号转导相关基因表达模式的改变。研究表明:经GA3处理的葡萄果实显著伸长,处理后14 d和成熟期葡萄果形指数分别为3.60和2.58,显著大于对照(2.09和1.81);同时,果实内细胞也显著伸长,处理后7 d和14 d细胞指数分别为2.38和2.68,显著大于对照(1.67和1.47);与对照组相比,果实中内源赤霉素、生长素和IAA-ASP的水平均显著升高;此外,GA3处理可显著提高葡萄果实中生长素合成和信号转导相关基因的表达水平。
关键词:‘金手指’葡萄;果实形状;GA3;生长素
赤霉素是果实发育过程中所需的主要激素之一,在植物从花到果的转变过程中起到了关键作用(陈杰忠&周碧燕,2002;丁长奎&章恢志,1988)其可以参与植物开花和坐果(Mariotti et al.,2011;Serrani et al.,2007)。GA3作为一种外源赤霉素,由于具有提高坐果率、增大果实和提高果实品质等方面的作用而被广泛应用于果树生产实践中。研究表明,GA3可以影响果实发育过程中的细胞分裂和细胞伸长(胡友军,2000)。此外,GA3还具有调运养分、吸引营养物质的作用(陶建敏 等,2003),可通过调节果实中营养物质的运输和分布来调控果实发育。说明其对果实形状建成具有显著的影响(程大伟 等,2021;Abu-Zahra,2010)。
已有大量研究表明GA3对于葡萄果形具有显著调节作用,Dokoozlian和Peacock(2001)发现GA3可以增加葡萄浆果长度;Casanova等(2009)在研究中进一步阐述了GA3对葡萄果实形状的影响主要是通过改变葡萄浆果细胞形态而实现的,GA3不影响浆果果皮细胞数,但增加果皮细胞直径;通过施用赤霉素生长抑制剂,如2-氯乙基三甲基氯化铵(CCC),还可以通过抑制GA3的合成从而使浆果变小(Coombe&Hale,1973);此外,在其他物种上也有许多GA3对果形具有显著调节作用的实例,在Chen等人(2020)的研究中分别对番茄施用GA3和赤霉素抑制剂PAC,结果表明GA3处理的番茄果实果形指数为1.25,显著高于对照(1.09),施用PAC使果形指数降至0.87,说明GA3可以显著提高番茄果形指数;在拟南芥和辣椒中,OFP 可能通过调节GA3含量变化来调节果形(Wang et al.,2007);草莓中发现了一种短果形的突变体sf,该突变体表现出GA3合成途径的缺乏(Wang et al.,2017);梁家伟(2014)和杜研等人(2013)的研究发现,在苹果果形发育过程中,果实中内源激素含量分布不均匀会导致苹果果实的偏斜,即非正常果形的苹果果实,但通过外源施用IAA、BA、GA3等植物生长调节剂,便可增大果形指数同时改善果实偏斜。Ahmed等人(2010)研究发现,在枣树上施用GA3,可以增大果长和果径。综上所述,GA3在调控果实形状方面具有重要的作用。
目前,关于外源施用GA3调控果实形状的研究多集中在果实生理特征上,分子层面上的研究还相对较少,为了阐明GA3在调控果实形状变化的内部分子机制,本研究通过制作石蜡切片、测定内源激素含量及检测激素信号通路相关基因的表达情况,从细胞及基因表达水平进行分析,为GA3调控葡萄果实形状提供理论依据。
1 材料方法
1.1 植物材料及其处理方法
试验于2020年在南京农业大学汤山葡萄教学试验基地进行,以‘金手指’葡萄为试验材料。采用平棚架避雨栽培,株行距为3.0 m×6.0 m。土肥水管理及病虫害防治同常规管理。以花序穗肩的第一个小穗上50%的小花开放为盛花期。于盛花期后3 d,选取三株树势健壮且相近的‘金手指’葡萄植株,使用25 mg/L的GA3溶液对花序进行处理,每个处理选取60穗左右,并做好标记,清水处理作为对照。分别于处理后0 h、6 h、12 h、24 h、48 h、4 d、7 d、14 d采集处理及对照的葡萄幼果并且采集成熟期葡萄果实。部分处理后6 h、24 h、48 h、4 d、7 d、14 d的样品置于FAA固定液中,4℃保存,用于后续石蜡切片的制作;部分处理后6 h、12 h、24 h、48 h、4 d、7 d的样品立即用液氮速冻,保存于-80℃冰箱中,用于后续总RNA的提取,其中,部分处理后48 h、4 d的样品用于后续内源GA3、生长素和IAA-ASP的测定。
1.2 葡萄表型观察
以处理组和对照组在处理后6 h、12 h、24 h、48 h、4 d、7 d、14 d时采集的果实为样品,使用体视显微镜进行观察。测量样品的纵径、横径并计算果形指数(果实纵径与果实横径的比值)。以处理组和对照组在成熟期的果实为样品。测量样品的纵径和横径并计算果形指数,处理组及对照组均含有20个样品。
1.3 细胞的形态观察
以处理组和对照组在6 h、24 h、48 h、4 d、7 d、14 d时采集的果实为样品制作石蜡切片。样品采集之后立即放在FAA固定液中进行固定,并置于4℃下进行保存。样品以乙醇/二甲苯系列脱水,石蜡包埋,切成3 µm厚的切片,干燥后用甲苯胺蓝进行染色。采用正置荧光显微镜显微成像系统观察细胞形态,测量细胞的纵径和横径并计算细胞指数(细胞纵径与细胞横径的比值),每次测量使用三个不同样品的三个切片(Cong et al.,2020)。
1.4 内源GA3、生长素和IAA-ASP的测定
以处理组和对照组在48 h和4 d时采集的果实为样品,采用高效液相色谱电喷雾串联质谱法(ESI-HPLC-MS/MS)测定样品内源GA3、IAA和IAA-ASP的含量。样品洗净后在液氮中研磨成粉末,称取0.5 g样品粉末,加入4 mL提取缓冲液(柠檬酸、2,6-二叔丁基-4-甲基苯酚和甲醇)。4°C振荡过夜后,10000 g离心15 min,取出上清液。沉淀反复浸提两次,合并所得上清液。在4°C摇动1 h,转速为120 rpm,随后超高速离心机10000 g离心15 min。然后将上清液在氮气下干燥,300 µL甲醇(0.1%甲烷酸)复溶,并通过0.22 µm的滤膜。色谱条件为:采用poroshell 120 SB-C18反相色谱柱(2.1 × 150, 2.7 um),进样量为2 µl。质谱条件为:喷雾电压4500 V,气幕、雾化器和辅助气体压力分别为15、65和70 psi,雾化温度为400°C。进行3次独立的生物重复试验(Cong et al.,2020)。
1.5 葡萄果实总RNA的提取
以处理组和对照组在6 h、12 h、24 h、48 h、4 d、7 d时采集的果实为样品,进行总RNA的提取,总RNA的提取采用多糖多酚植物总RNA提取试剂盒(TIANGEN, China),按照说明书进行操作。取2 μL RNA样品进行琼脂糖凝胶电泳,对RNA样品的质量进行检测。用Nanodrop ND-1000 Spectrophotometer 检测RNA浓度。
1.6 荧光定量PCR
使用实时荧光定量PCR仪-Q5进行qRT-PCR分析,反应体系按SYBR Green PCR Master Mix(Takara)荧光定量qPCR试剂盒说明书进行。使用snapgene软件设计荧光引物,引物见表2-1,VvUbiquitin作为内参基因。进行3个生物学重复,使用2-△△Ct公式计算相对表达量(Livak&Schmittgen,2002)。
1.7 统计分析
用Excel 2010和SPSS 17.0进行数据处理,Adobe photoshop CC 2019作图,SPSS进行显著性分析。
2 结果分析
2.1 GA3处理葡萄果实形态观察
2.1.1 葡萄幼果形态观察
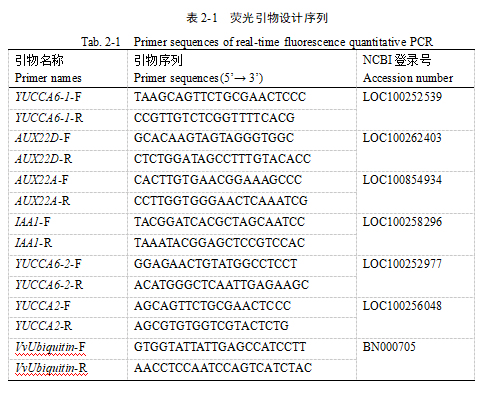
葡萄幼果形态自GA3处理后48 h开始出现差异(图2-1 A),处理组果实明显伸长,并且随着时间的变化处理组与对照组之间的差异越来越明显。幼果纵横径测量结果显示,GA3处理后0 h-24 h,幼果的纵径和横径均没有发生明显的改变(图2-1 B, C);GA3处理后48 h幼果的纵径开始增大,至GA3处理后14 d时处理组的果实纵径约为对照组的1.56倍(图2-1 B),但果实横径却没有表现出相似的变化规律,处理组幼果的果实横径较对照组均无显著差异(图2-1 C)。计算果形指数可知,自GA3处理后4 d起,处理组与对照组果形指数开始出现显著性差异,处理后4 d处理组果形指数为2,对照组为1.74,处理后7 d处理组果形指数为3.12,对照组为1.83,处理后14 d处理组果形指数为3.6,对照组为2.1(图2-1 D),果形指数均显著增大且处理组与对照组之间的差异越来越大。总而言之,处理组果实横径大体上与对照组相近,并没有表现出明显变化,但处理组果实纵径自GA3处理后48 h开始表现出稳定增长的趋势,其与对照组之间的差异越来越大,因此,处理组与对照组果形指数之间的差异也越来越显著,至GA3处理后14 d处理组幼果的果形指数达到3.6,约为对照组的1.7倍。说明GA3处理可使‘金手指’葡萄幼果伸长,可初步推测GA3处理后24 h-48 h之间葡萄幼果形态开始发生改变,GA3处理48 h之后葡萄幼果开始显著伸长。
2.1.2 葡萄成熟期形态观察
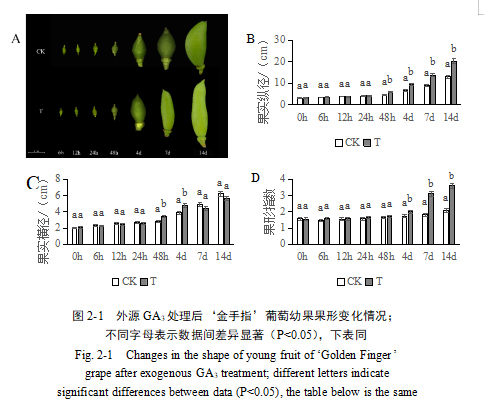
‘金手指’葡萄果实成熟后,处理组与对照组葡萄外观出现了明显的变化。与对照组相比,GA3处理的葡萄果粒伸长,整体串型更为松散,葡萄果穗变长,葡萄的生长空间更大(图2-2 A);单粒果果实形状较对照组明显伸长(图2-2 B),单粒果的纵径变大(图2-2 C),但横径不发生明显改变(图2-2 D),处理后果形指数为2.58,显著大于对照(1.81)(图2-2 E)。结果表明,适宜浓度的GA3处理可以显著促进葡萄果实伸长,增大葡萄果粒果形指数。此外,GA3处理对葡萄果穗具有一定的拉长作用。
2.2 GA3处理葡萄果实细胞形态观察
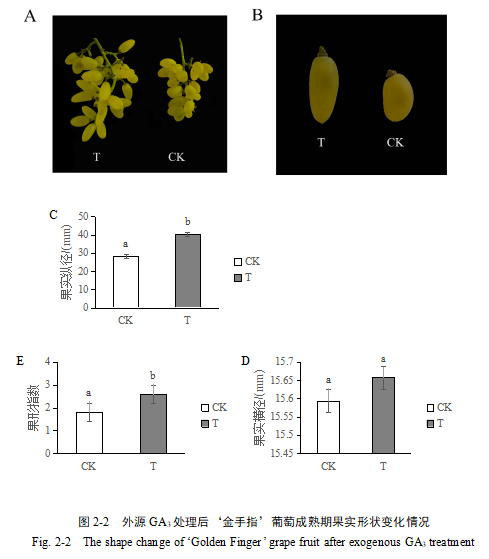
为进一步观察GA3处理对细胞发育的影响,以GA3处理后6 h、24 h、48 h、4 d、7 d、14 d的‘金手指’葡萄果实为样品制备石蜡切片。结果显示,GA3处理的葡萄果实细胞大小明显大于对照组,且处理组细胞较对照组明显伸长(图2-3 A, B);相同的视野内处理组所能看到的细胞个数明显少于对照组,说明处理后葡萄果实内部细胞面积增大。细胞纵横径测量结果显示,GA3处理后6 h细胞纵径和细胞横径均无明显变化(图2-3 C, D);GA3处理后24 h细胞纵径开始增大,至GA3处理后14 d时处理组的细胞纵径约为对照组的2倍(图2-3 C),但细胞横径却没有表现出相似的变化规律,处理组细胞横径较对照组均无显著差异(图2-3 D)。计算细胞指数可知,自GA3处理后48 h起,处理组与对照组细胞指数开始出现显著性差异,到GA3处理后14 d时处理组的细胞指数达到了2.68,但对照组仅为1.47(图2-3 E)。总而言之,处理组细胞横径大体上与对照组相近,并没有表现出明显变化,但处理组细胞纵径自GA3处理后24 h开始表现出稳定增长的趋势,这也导致了处理组与对照组细胞指数差异越来越显著。这些结果表明,GA3在果实发育早期通过促进细胞膨胀和伸长来调控果实大小,起到促进果实的伸长的效果,且GA3促进细胞的伸长的作用可能开始于处理后24 h。
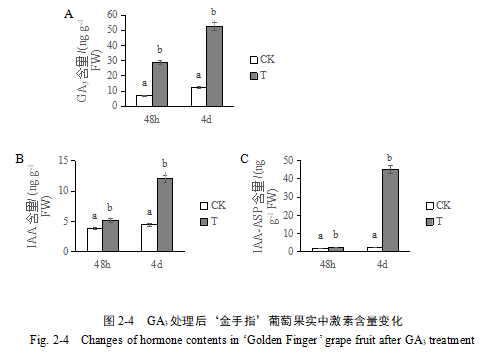
2.3 GA3处理对‘金手指’葡萄果实中内源赤霉素、生长素和IAA-ASP的水平的影响
通过ESI-HPLC-MS/MS方法测定了GA3处理和清水对照样品中内源赤霉素、生长素和IAA-ASP的含量。结果表明,GA3处理之后48 h和4 d后,处理组葡萄果实中内源赤霉素含量分别为对照组的4.2倍和4.3倍(图2-4 A);GA3处理48 h后,葡萄果实中IAA和IAA-ASP含量开始较对照组升高,分别为对照组的1.32倍和1.37倍,处理后4 d处理组和对照组IAA和IAA-ASP含量差异显著增大,分别为对照组的2.7倍和18.3倍(图2-4 B, C)。说明外源GA3处理会使‘金手指’葡萄果实中的内源赤霉素、IAA和IAA-ASP含量升高且处理组和对照组含量在处理后48 h差异较小,处理后4 d差异更为显著。
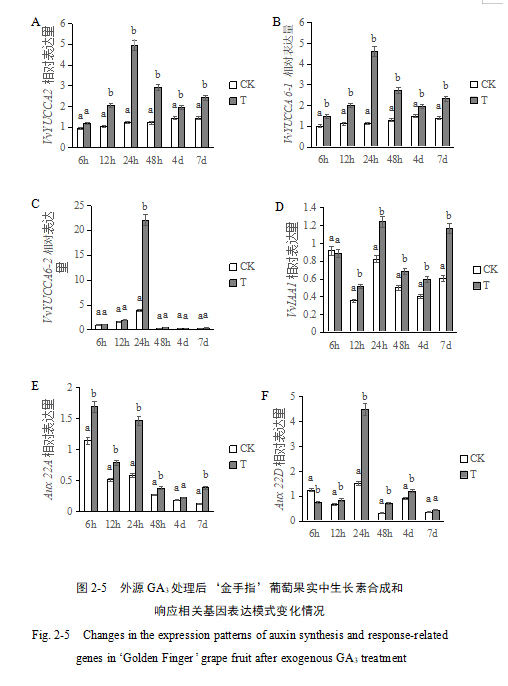
2.4 GA3处理对‘金手指’葡萄果实中生长素生物合成和信号转导相关基因表达模式变化影响
为了进一步探究生长素对果实形状的影响,本研究测定了葡萄果实中生长素生物合成和信号转导相关基因的表达。YUCCA是生长素合成过程中的关键基因,挑选了三个YUCCA基因家族的成员VvYUCCA2、VvYUCCA6-1及VvYUCCA6-2进行qRT-PCR检验。qRT-PCR结果显示,与对照组相比,‘金手指’葡萄中生长素合成相关基因VvYUCCA呈升高趋势,在处理后24 h时达到最高且与对照组含量差异最大。三个基因表达趋势较为一致。GA3处理后24 h时,‘金手指’葡萄中VvYUCCA2、VvYUCCA6-1含量为对照组的4-5倍,并随着时间的推移与对照之间的差异逐渐减小,表明GA3处理可以显著提高‘金手指’葡萄中的VvYUCCA2、VvYUCCA6-1及VvYUCCA6-2基因的表达水平(图2-5 A, B, C)。IAA1是生长素/吲哚-3-乙酸家族成员,是一种生长素信号转录调控蛋白,与对照组相比,VvIAA1在处理6 h后无明显差异,在处理12 h-7 d后,表达水平显著升高。说明GA3处理可以显著提高‘金手指’葡萄中IAA1基因的表达水平(图2-5 D)。Aux22A和Aux22D是两个生长素信号转导相关基因,GA3处理后Aux22A的表达量呈升高的趋势,然而Aux22D却表现出先下降后升高的趋势,但整体而言,GA3处理可以显著上调这两个基因的表达。Aux22A和Aux22D处理组与对照组差异最大的时期都为处理后24 h,此趋势与三个VvYUCCA基因一致(图2-5 E, F)。
3 讨论
GA3对果实形状具有显著调节作用,可以增大果实纵径和果形指数,从而促进果实伸长(娄玉穗 等,2019;谢周 等,2010)。胡友军(2000)的研究结果表明GA3促进果实伸长作用可能是通过影响果实内部细胞形态变化而实现的,同时,Lucas等人(2008)发现GA3具有促进细胞伸长和膨胀的作用。在本研究中发现外源GA3处理葡萄果实后,果实形态明显改变且果实纵径和果形指数显著变大;果实内部细胞明显变大且细胞纵径和细胞指数显著变大,这与前人的研究一致。此外,GA3处理后的葡萄果穗也有一定程度的伸长,这可能与细胞的伸长有关。以上结果可初步推测出GA3是通过影响果实内部细胞的形态进而影响果实形状。
施用外源植物生长调节剂能够影响植物内源激素的水平(陈瑶 等,2020)。已有研究表明外源施用GA3可以提高内源赤霉素的含量(吴月燕 等,2011);吴建明等(2009)在研究中也阐述了利用外源GA3处理甘蔗幼茎可提高幼茎内源赤霉素和生长素含量。本研究结果表明:GA3处理后葡萄幼果中内源赤霉素含量升高,这与陈显(2013)的研究结果相似;处理后内源IAA含量显著上升,这与黎正英等人(2021)的结果相似。此外,IAA-ASP是生长素的主要储存形式(Hayashi et al.,2021),其含量显著升高可表明果实内IAA含量变化较大,因此可推测外源施用GA3可能对内源生长素的水平具有显著影响。
YUCCA作为生长素合成过程中的关键基因,其被激活或者过表达都会导致游离的生长素含量升高,且YUCCA基因家族功能冗余,可能需多个基因协同发挥作用(Kim et al.,2013)。本研究中三个YUCCA基因的表达量变化趋势基本一致,GA3处理后其在果实内部的表达水平均升高,说明GA3处理会影响果实内部YUCCA基因的表达;IAA1编码的IAA1蛋白是Aux/IAA蛋白家族的成员,该家族成员可响应生长素(Yang et al.,2004)。在本研究中GA3处理后果实内IAA1的表达水平显著上升,说明GA3处理会影响果实内部IAA1基因的表达。此外,处理后果实内Aux22A呈升高的趋势,Aux22D虽表现出先下降后升高的趋势,但两者整体表达量变化趋势较为一致,处理组表达水平均较对照组高。根据以上结果可推测外源施用GA3可影响果实内源生长素的合成和信号转导。
综上所述,在‘金手指’葡萄盛花后3 d施用外源GA3可促进果实伸长,且该结果可能是由于内部细胞伸长所导致的。此外,外源GA3可能对内源GA3和IAA水平产生影响且其调控葡萄果实形状变化的机制可能与GA3以及IAA的合成代谢和信号转导相关。