北京综合试验站
一 试验材料和测定项目
供试葡萄接穗为欧亚种(Vitis vinifera L.)鲜食品种‘瑞都香玉’(果皮黄绿色,XY)和‘瑞都红玉’(果皮红紫色,HY),砧木品种分别为河岸葡萄(V. riparia Michixi)× 沙地葡萄(V. rupestris Scheele)组合选育的‘3309M’,冬葡萄(V. berlandieri Planchon)× 沙地葡萄(V. rupestris Scheele)组合选育的‘1103P’和‘110R’,冬葡萄(V. berlandieri Planchon)× 河岸葡萄(V. riparia Michixi)组合选育的‘5BB’和‘SO4’。以‘瑞都香玉’和‘瑞都红玉’自根苗作为对照。共计12个处理。随机区组试验,每小区4株,选取长势基本一致的3株树,3个重复。2018年5月11日(初花期)、6月6日(新梢生长期)、7月18日(果实转色期)和8月15日(果实成熟期)分别采集新梢基部向上第3 ~ 4节叶片以及新梢梢尖5 ~ 6 cm处组织,液氮保存,–80 ℃超低温冰箱中保存。2018年8月底和2019年8月底,每个处理于树体东、西两侧各部位随机采取6 ~ 8穗果实带回实验室进行品质鉴定。采用酶联免疫分析法测定样品中吲哚乙酸(IAA)、脱落酸(ABA)、赤霉素(GA3)和玉米素核苷(ZR)含量。
用Microsoft Excel 2007软件整理、统计数据以及作图,用Sigmaplot 12.0数据分析软件进行单因素方差(ANOVA)分析以及作图,Student-Newman-Keuls法多重比较(P < 0.05)。
二 结果与分析
2.1 ZR
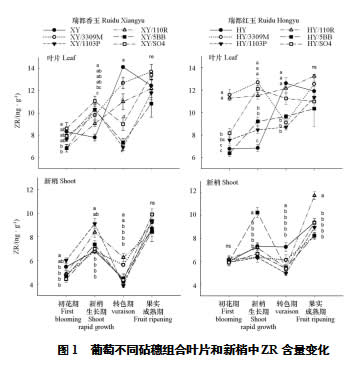
对不同砧穗组合的叶片和新梢中ZR含量进行测定。结果(图1)显示自根苗叶片ZR在生长季内的变化趋势与嫁接苗不同:除瑞都红玉/3309M以外,其余嫁接苗组合在初花期的ZR含量为全年最低,新梢生长期显著升高,进入转色期后不同组合表现不同,到果实成熟期所有组合又稍有升高;自根苗则表现为在新梢生长期的叶片ZR不高于初花期,但是进入转色期后有大幅度升高,达到峰值,到果实成熟期后有所下降,但仍显著高于新梢生长期。长势较旺的是以110R、SO4和5BB为砧木的组合,其中SO4组合在新梢生长期ZR处于较高水平,5BB组合在转色期处于较低水平,110R与两种接穗品种的组合表现出了差异:瑞都香玉/110R处于中等水平,瑞都红玉/110R处于较高水平,这与2018年和2019年110R组合接穗品种基部粗度最大相对应。长势较弱的3309M组合的ZR含量全年都处于较高水平,1103P处于低水平。
新梢的ZR含量小于叶片,各组合在生长季内变化趋势一致:新梢生长期和果实成熟期高于初花期和转色期。新梢生长期XY/1103P、XY/110R和HY/5BB含量高于其他组合;转色期XY/110R、XY/3309M和HY显著高于其他组合。
2.2 IAA
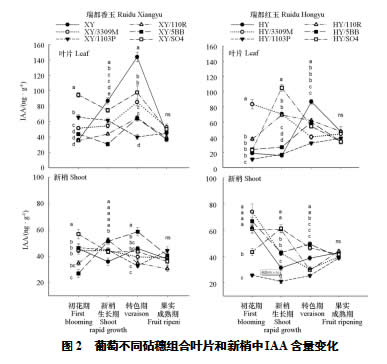
葡萄幼树体内含有大量IAA,其含量远高于ZR。叶片IAA结果(图2)表明,瑞都香玉自根苗在初花期含量低,但是新梢生长期和转色期含量迅速升高,显著高于其他组合,到果实成熟期后降至初花期的水平;瑞都红玉自根苗的峰值同样在转色期,与瑞都香玉不同的是瑞都红玉在新梢生长期的含量显著低于其他组合。与ZR结果类似,生长季内SO4组合的IAA含量高于其他组合:瑞都香玉/SO4在初花期、新梢生长期和转色期的含量显著高于其他组合;瑞都红玉/SO4在初花期显著低于瑞都红玉/3309M,但在新梢生长期达到峰值,显著高于其他组合,到转色期后与瑞都红玉/110R和瑞都红玉/5BB处于同一水平,高于瑞都红玉/1103P和瑞都红玉/3309M。瑞都香玉/5BB和瑞都香玉/110R全年处于较低水平,瑞都红玉/110R在新梢生长期和转色期的IAA含量较高,瑞都红玉/5BB在新梢生长期含量低,转色期含量有所提高。
新梢内IAA含量远低于叶片。初花期时各组合之间差异显著,瑞都香玉/SO4、瑞都红玉/3309M、瑞都红玉/5BB、瑞都红玉和瑞都红玉/110R处于较高水平,瑞都香玉/5BB和瑞都红玉/1103P最低。新梢生长期,瑞都香玉组合差异不显著;瑞都红玉/SO4和瑞都红玉/110R含量最高,瑞都红玉/1103P最低。转色期,瑞都香玉/5BB、瑞都红玉/5BB和瑞都红玉/SO4含量最高;1103P和110R组合最低。
2.3 GA3
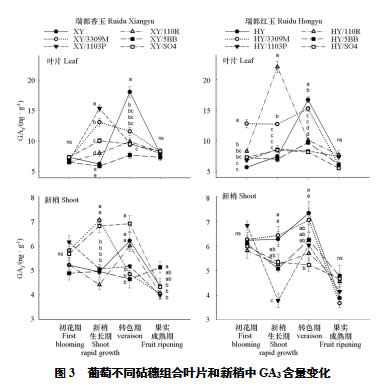
2个接穗品种自根苗的叶片GA3含量在转色期达到顶峰,显著高于其余砧穗组合,在其他3个时期则处于低水平(图3)。新梢生长期,瑞都红玉/110R含量最高,其次是瑞都香玉/1103P、瑞都香玉/3309M和瑞都红玉/33090M,含量最低的是瑞都香玉/5BB和瑞都红玉/5BB。转色期,瑞都香玉/3309M和瑞都红玉/3309M在砧穗组合中含量最高,瑞都香玉/5BB、瑞都红玉/5BB和瑞都红玉/ 1103P含量最低,其余组合处于中间水平。综合而言,以3309M为砧木的组合叶片GA3处于较高水平,而长势强旺的SO4和5BB组合则处于中低等水平。
新梢中的GA3含量远小于叶片。与ZR和IAA相似,各组合之间的差异主要体现在新梢生长期和转色期。新梢生长期瑞都香玉/3309M、瑞都香玉/SO4、瑞都红玉/3309M和瑞都红玉的含量高于其他处理,瑞都红玉/1103P最低。转色期瑞都香玉/SO4、瑞都香玉、瑞都香玉/110R、瑞都红玉和瑞都红玉/3309M的含量高于其他处理。果实成熟期的GA3为全年最低,瑞都香玉/5BB此时期含量最高。
综合ZR、IAA和GA3这3种促生长类激素结果可知,在6月初的新梢生长期和7月中旬的转色期是激素含量高峰期,各组合在这2个时期的激素含量差异明显,以110R和SO4为砧木的组合叶片和新梢激素含量值常处于较高水平,以5BB为砧木的组合叶片含量低,但是新梢含量高。瑞都红玉/3309M叶片的促生长类激素在初花期的含量均显著高于其他组合,其当年的可溶性固形物也较高(表1),说明3309M促进了瑞都红玉的发育进程,但是其促生长类激素在新梢生长期和转色期有所下降,导致瑞都红玉/3309M的长势不强于其他组合。
2.4 ABA
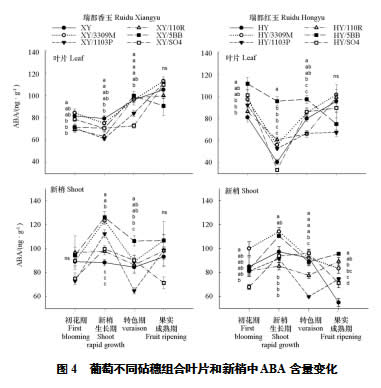
从初花期到新梢生长期,叶片中抑制类激素ABA的含量减小;从新梢生长期到转色期,再到果实成熟期,除瑞都红玉/5BB之外,所有组合的ABA含量一直增加(图4)。瑞都香玉各组合差异不大;瑞都红玉各组合之间差异则非常显著,瑞都红玉/5BB的变化趋势与其他组合不同,其在新梢生长期的ABA含量非常高,到果实转色期后含量却显著下降。此外,瑞都红玉组合在新梢生长期的ABA含量低于瑞都香玉组合,其中以瑞都红玉/SO4含量最低,与其长势强旺相吻合。
新梢的变化趋势与叶片不同,大多数组合从初花期到新梢生长期ABA升高,到转色期下降,进入果实成熟期后又升高。以3309M和5BB为砧木的组合ABA全年多处于较高水平,对其长势有一定抑制作用。表现突出的是瑞都香玉/SO4、瑞都红玉/SO4和红玉自根苗,这3个处理从转色期到果实成熟期的ABA不升反降,有利于新梢在第二次生长高峰期的生长。
三 结论与讨论
本研究中测定了各砧穗组合的ZR、IAA、GA3和ABA在初花期、新梢生长期、果实转色期和果实成熟期的动态含量变化,发现在6月初的新梢生长期和7月中旬的果实转色期叶片和新梢促生长类激素含量较初花期和果实成熟期高,其中以110R和SO4为砧木的组合叶片和新梢激素含量常处于较高水平,以5BB为砧木的组合叶片含量低,但是新梢含量高。由于瑞都红玉为早熟品种,瑞都香玉为中熟品种,因此瑞都红玉的促生长类激素含量的峰值多出现在新梢生长期,瑞都香玉的峰值在转色期,而且瑞都红玉组合在新梢生长期的ABA含量低于瑞都香玉组合,与品种特性吻合。以SO4为砧木的组合还有1个特点:新梢ABA含量从转色期到果实成熟期不升反降,有利于新梢在第2次生长高峰期内的生长。