太谷综合试验站
赵旗峰 黄丽萍 王敏 刘晓婷 张晓萍 马小河
摘要:【目的】对山西太谷国家葡萄种质资源圃内保存的资源,通过表型性状鉴定分析筛选出不同葡萄种质资源进行SSR分析,揭示这些不同来源的葡萄种质资源之间亲缘关系和群体遗传结构,为葡萄种质资源的科学管理和分子标记辅助育种提供参考。【方法】以100份葡萄种质为材料,依据基因组DNA测序挖掘出的SSR引物数据,筛选出33对核心多态引物,根据标准分子量记录扩增DNA多态性条带,获得数据矩阵。利用每对SSR引物在100份葡萄种质资源中等位基因组成计算33对SSR引物的位点多态性信息含量(PIC)、100份葡萄种质资源个体间的Nei’s遗传距离,并利用MEGA7.0软件构建NJ邻接聚类树并推断葡萄原始集合与核心集合的遗传结构。【结果】100个样本中的33对SSR标记共检测到423个等位基因,每对引物扩增条带数4~26条,平均扩增条带数为12.82条。葡萄种质资源之间的Nei’s遗传距离约为0.4~0.8,占资源总量的97.80%。通过遗传结构分析并预测其最佳分组数K为2,即葡萄核心集合主要分为两个亚群,分别为欧亚种群的鲜食葡萄和杂种群,其中杂种群涵盖了欧美杂种、美洲种、中国野生种及欧亚种的酿酒品种。来自国外的美洲种的砧木和来自山西的野葡萄聚在一起,同时与欧美杂种和酿酒品种也聚为一类。可能是由于在人为选种和选用目的的不同造成各品种之间遗传背景的混杂。这一结论还需进一步研究证明。【结论】通过对基因型遗传关系和基因结构分析,为野生和栽培品种间遗传关系的分析提供了新的证据。
关键词:葡萄;SSR分子标记;亲缘关系;遗传结构
葡萄属于葡萄科(Vitaceae)葡萄属(Vitis L.),是栽培历史最悠久的植物之一。中国的葡萄栽培约始于2000多年前的汉武帝时期,是葡萄属植物的原产国之一。据不完全统计,目前我国入圃保存的葡萄种质资源约有3000份,品种繁多,种质资源、基因资源极为丰富,对这些资源进行多样性评价,可以有效的保护植物遗传多样性,并为培育不同特性的新品种提供基本材料;另外,由于定向培育新品种使种间缺乏基因流动,遗传背景越来越狭窄,新品种培育和抵御病虫害等方面都有较大的弊端。因而深入地利用各种方法特别是现代分子生物学技术来研究作物遗传多样性,对提高葡萄育种、种质资源管理及其保护水平,具有重要的理论意义和实际意义。太谷国家葡萄种质资源圃目前保存了17个种669份葡萄种质资源,为了加快对圃内保存资源的开发利用研究,对圃内保存的资源进行亲缘关系和群体遗传结构的研究意义重大,同时为葡萄核心种质资源的构建和遗传育种等提供支撑。
DNA标记技术被广泛应用于种质资源遗传多样性研究。其中SSR标记技术以其不受环境、发育时期、不同器官等限制的优越性决定了它具有更高的可靠性和高效性,更容易从分子水平上去研究物种的亲缘关系、种质资源保存、构建图谱及辅助育种等,已成为当前品种鉴定的主要技术。同时SSR标记在葡萄种质资源鉴别、揭示和区分同名异物和同物异名等方面已经得到广泛应用。Thomas和Scoot等研究认为,SSR标记可以把葡萄种、种间杂交品种和纯种品种正确区分。Botta等的研究结果也证明,利用SSR标记对葡萄种质资源和品系遗传多样性进行分析,具有较高的可靠性和现实意义。Martínez等利用SSR技术分析了南美葡萄品种的遗传多样性;温景辉、方连玉等利用SSR标记对葡萄种质进行了多态性分析,成功的将山葡萄品种、欧亚种及美洲杂种区分开。吴子龙等利用SSR标记对8个山葡萄及山欧杂种葡萄品种进行了区分。邹瑜等利用SSP技术对70份毛葡萄种质资源进行了多样性分析,阐明了毛葡萄种质的亲缘关系。郭春苗等利用SSP技术对葡萄品种(系)遗传多样性进行了分析,在一定程度上反映了品种之间的亲缘关系。VVS2、VVMD5、VVMD7、VVMD25、VVMD27、VVMD28、VVMD32、VrZAG79、VrZAG62这9对SSR引物已作为国际上葡萄品种鉴定的通用标记。尹玲等、成冰、杨航宇等、李贝贝等也利用国际通用的 SSR 标记分别构建了不同份数的葡萄品种遗传图谱。王富强、李贝贝等从 137 个葡萄标记中筛选出 30 个标记,构建了一套适用于中国的葡萄 SSR 分子标记体系,同时为保证鉴定效率和国际品种鉴定接轨 ,选择 VVMD28、VVMD32、VVMD27、VrZAG79、VVMD7、VrZAG62、VVMD25、VVS2、VVMD5 这 9 个标记作为体系的核心引物。为了对国家果树种质资源太谷葡萄圃保存的种质资源进行深入鉴定评价,本研究利用SSR分子标记对100份葡萄种质资源进行遗传多样性分析,揭示这些不同来源的葡萄种质资源之间的亲缘关系和群体遗传结构,为葡萄种质资源的科学管理和分子标记辅助育种提供参考。
1 材料与方法
1.1 材料
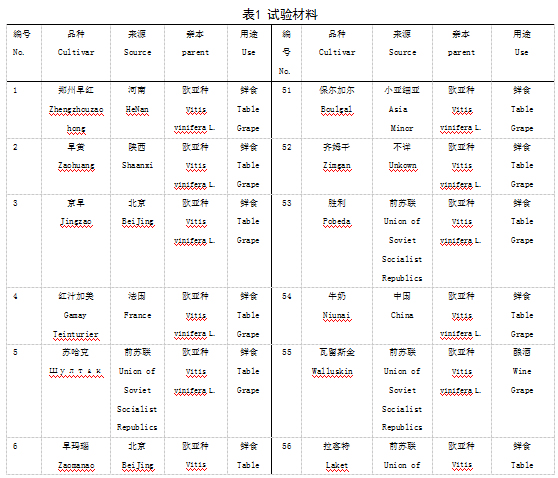
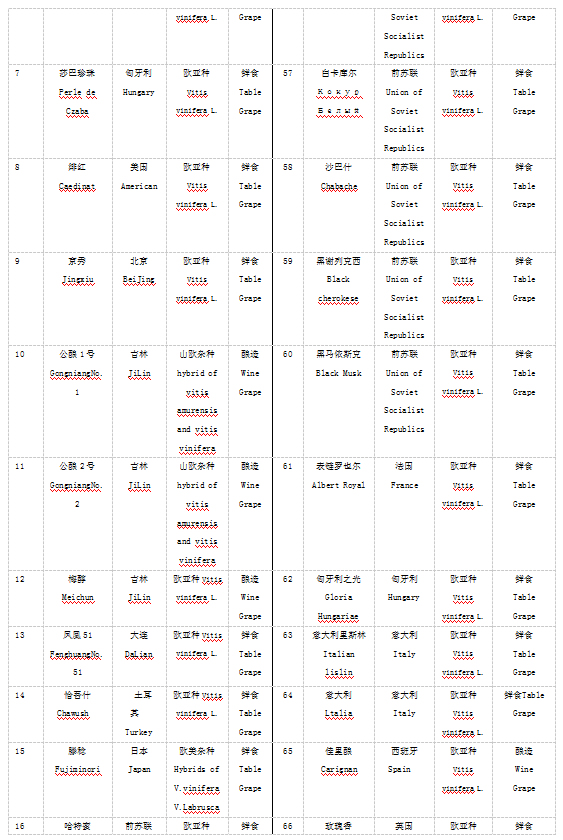
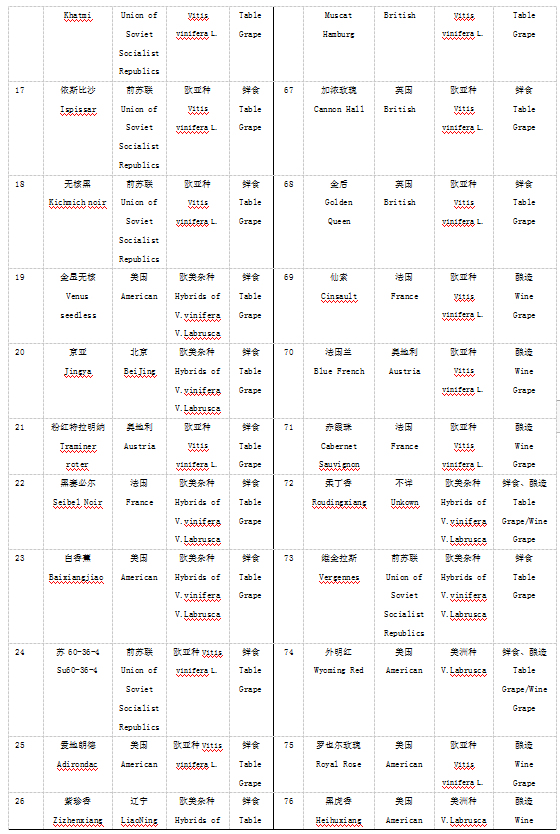
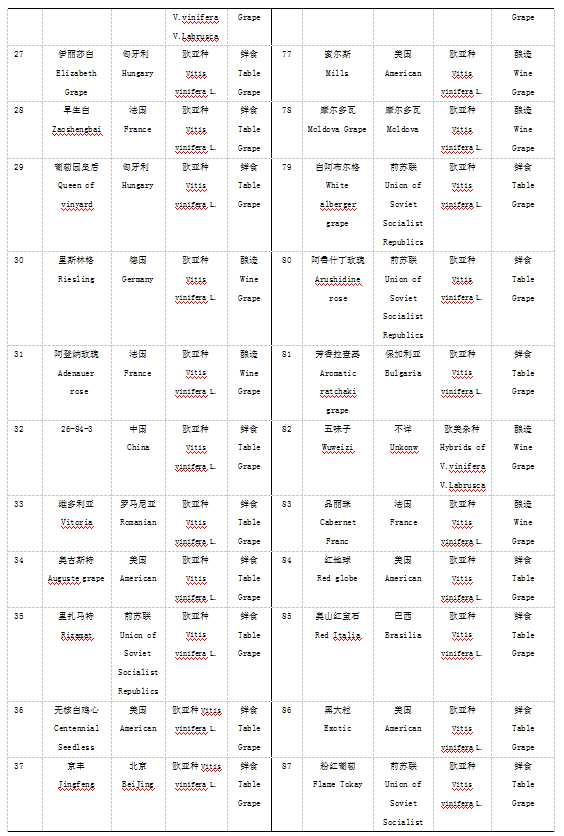
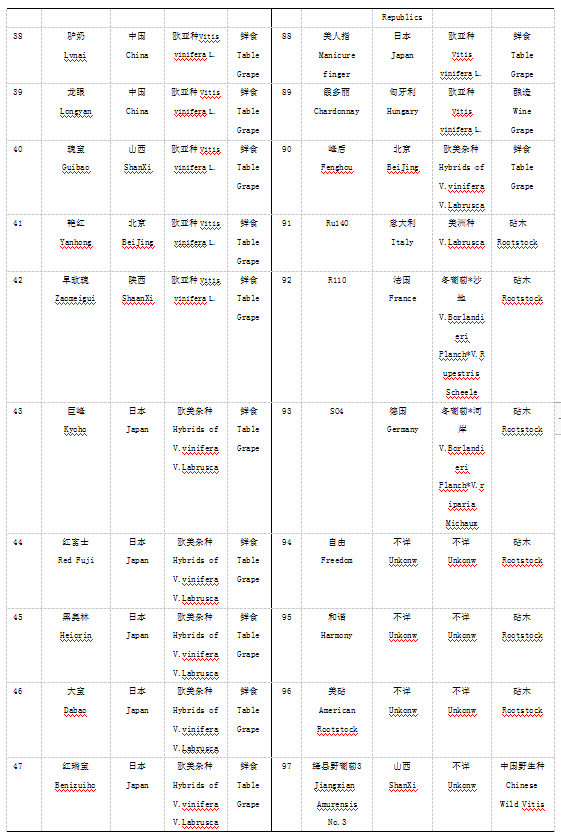
本实验材料根据表型特征从国家果树种质资源太谷葡萄圃保存的资源中选取了100份葡萄种质资源,2016年采集葡萄幼嫩叶片,液氮处理后-70℃保存备用。具体名称见表1.

1.2 试验方法
1.2.1 基因组DNA提取
取新鲜叶片在液氮中充分研磨成粉末,并转移至1.5 ml的离心管中,加入600 ul 65℃预热的Buffer PCB和12 ul β-巯基乙醇,震荡混匀,置于65℃水浴25 min,加入等体积的氯仿,充分混匀,12000 rpm离心5 min。吸取上层水相至干净的1.5 ml的离心管中,加入等体积Buffer BD,颠倒混匀3-5次,再加等体积的无水乙醇,充分混匀后用移液器将其全部加入到吸附柱中,室温静止2 min,10000 rpm,离心1 min,倒掉收集管中废液,将吸附柱放回收集管中,加入500 ul Wash Solution,10000 rpm,离心1 min,倒掉收集管中废液,将吸附柱放回收集管中,12000 rpm,离心2 min,取出吸附柱,放入一个新的1.5 ml离心管中,在吸附膜中央加入50ul TE Buffer,静止3 min,12000 rpm,离心2 min,得到DNA溶液置于-20 ℃保存。
1.2.2 SSR-PCR扩增及电泳
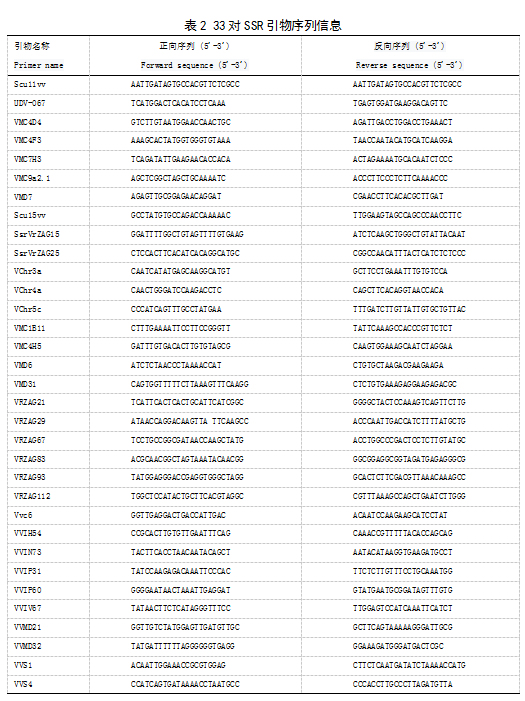
本试验以前期葡萄基因组测序挖掘出的SSR引物为基础,从中筛选出了33对核心多态引物(表2)。SSR-PCR扩增在3730XL测序列分析仪上进行。PCR反应体系为25 μL,其中包括1 μl模板DNA,0.2 μL5U/μl TaKaRaTaq酶,2.5 μlTaq buffer,2.0 μL25m mol/LMgcl2,0.5 μl 10m mol/L dNTP,0.5 μl引物F,0.5 μl引物R,17.8 μlH2O。PCR反应程序为:95 ℃预变性3 min;95 ℃变性30 s,60 ℃退火30 s,72 ℃延伸30 s,10 c个循环;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,20 c个循环,72 ℃修复延伸6 min。扩增产物进行Fragment AnalyzerTM全自动毛细管电泳检测。
1.2.3 数据统计分析
根据标准分子量记录扩增DNA多态性条带(片段),获得数据矩阵。利用Gen AlEx6.503软件计算等位基因数Na(Number of alleles,Na),有效等位基因数(Effective number of alleles,Ne)、Shannon’s信息指数(Shannon’s information index,I)、期望杂合度He(Expected heterozygosity,He)、观测杂合度Ho(Observed heterozygosity,Ho)、Nei’s遗传多样性H(Nei’s gene diversity index,H),并统计每对SSR引物在100份葡萄种质资源中等位基因组成,包括稀有等位基因数(≤1%)、共有等位基因数(1%–≤20%)、最常见的等位基因数(>20%)。利用Power Marker软件计算33对SSR引物的位点多态性信息含量(PIC)、100份葡萄个体间的Nei’s遗传距离,并利用MEGA7.0软件基于Nei’s遗传距离构建100份葡萄种质资源的NJ邻接聚类树;利用STRUCTURE软件基于贝叶斯聚类分析方法推断葡萄原始集合与核心集合的遗传结构,其中群体数目(K)设为1~10,对每个K值模拟运算10次,设不作数迭代(lengthofburn-inperiod)开始时的马尔科夫链蒙特卡洛(MCMC)为100000次,不作数迭代后的MCMC为1000000次,最后将Structure结果导入在线网站Structure Harvester预测确定最佳的K值(分组数)。
2 结果与分析
2.1 33对SSR引物的等位基因组成
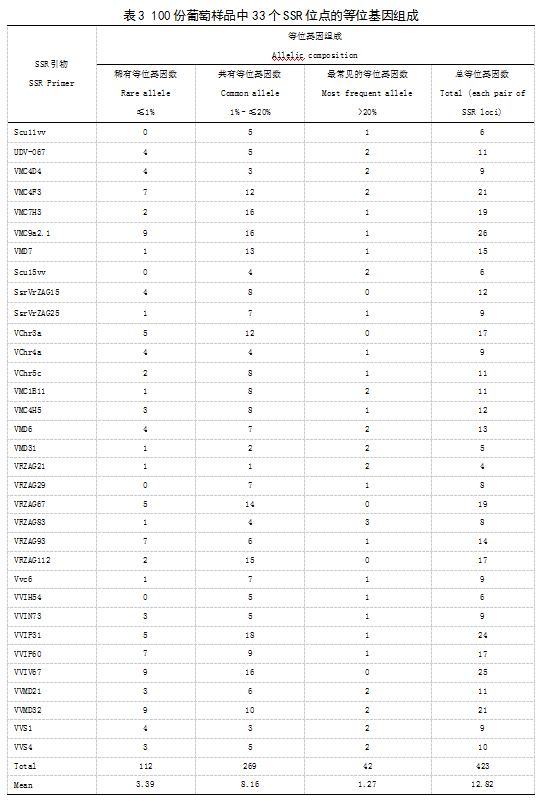
本实验利用33对SSR引物在100份葡萄种质资源中共检测到423个等位基因,每对引物扩增条带数4~26条,平均每对引物扩增条带数为12.82条。33对SSR引物在100份葡萄资源中扩增的等位基因组成中稀有等位基因数(≦1%)范围在0~9个,平均等位基因数有3.39个,共有等位基因(1~≦20%)为8.26个,最常见的等位基因数(>20%)为1.67个(表3)。
2.2 SSR引物多态性分析
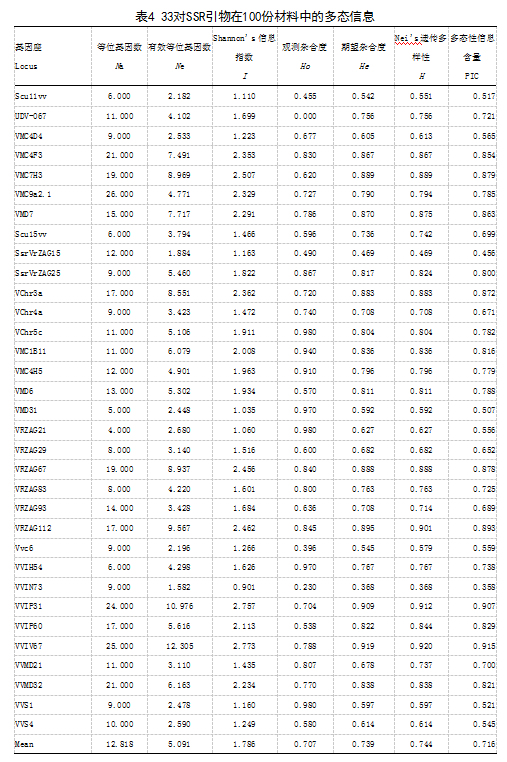
从70对SSR引物中筛选出33对能扩增出清晰稳定条带的引物(表4)。结果表明33对引物均能扩增出稳定清晰的DNA条带(表4)。33对SSR引物共扩增出351条DNA条带,多态性条带比率(PPB)为99.39%,每对引物扩增条带数4~21条,平均每对引物扩增条带数为10.64条,扩增条带大小范围为87~406bp。所有引物中,编号VMC9a2.1的引物扩增条带数最多,为21条,在引物评价种质资源遗传多样性的方法中,扩增条带数越多说明引物鉴别力越强,因此,VMC9a2.1为鉴别能力较强的引物,可作为种群内多样性鉴别的首选引物。检测到有效等位基因数(Ne)变化范围是1.582-12.305,平均为5.091;位点多态性信息含量(PIC);根据BOTSTEIN等提出衡量基因变异程度高低的多态性信息量指标PIC;PIC>0.5时,该引物为高度多态性信息引物;0.25<PIC<0.5,为中度多态性信息引物;PIC<0.25,为低度多态性信息引物。本研究中PIC变幅为在0.358-0.915之间,平均为0.716,均为高度或重度多态性信息引物。基因多样性(H)变化范围为0.368-0.920,平均为0.744。
2.3 100份葡萄种质遗传距结构分析
2.3.1 遗传距离及聚类分析
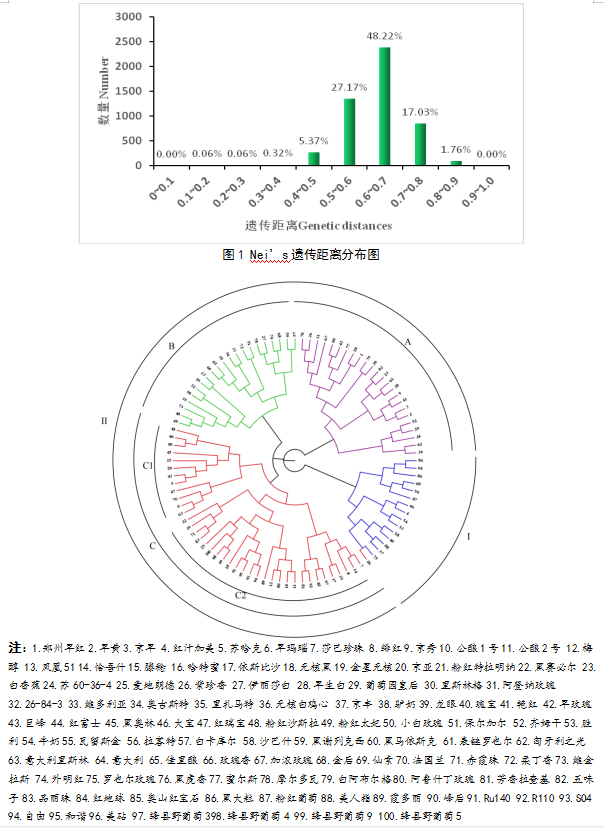
本实验利用对100份葡萄种质资源进行Nei’s遗传距离分析,结果得出:约为0.4~0.8,占种质资源总量的97.80%(图1)。48.22%在0.6~0.7之间,27.17%在0.5~0.6之间。17.03%在0.7~0.8。利用MEGA7.0软件基于遗传距离建立NJ遗传聚类树(图2),结果得到100份葡萄共分为2组,第一组(I组)基本为我国自育品种或亲本为我国古老品种的欧亚种的鲜食葡萄资源群;第二组(II组)较为复杂,可分为A,B,C三个亚组,其中A亚组多为来源于苏联及亚美尼亚高加索等地的欧亚种鲜食资源群,B亚组多为来源于欧洲大陆的欧亚种鲜食兼用葡萄资源群,C亚组又可分为C1,C2两个次亚组,C1次亚组主要为欧美种群鲜食葡萄资源,C2次亚组主要为欧亚种酿酒葡萄资源及野生砧木资源群。聚类结果表明,(1)分子标记分析的亲缘关系与其遗传背景具有显著相关性,鲜食,酿酒,砧木等用途不能成为区别的有效依据;(2)亲缘关系与亲本有一定的关系,但取决于其遗传力,SSR能比较准确地检测基因型之间的遗传背景与遗传关系;(3)欧美杂种与欧亚酿酒葡萄品种及野生种质资源亲缘关系较近。

2.3.2 主坐标分析
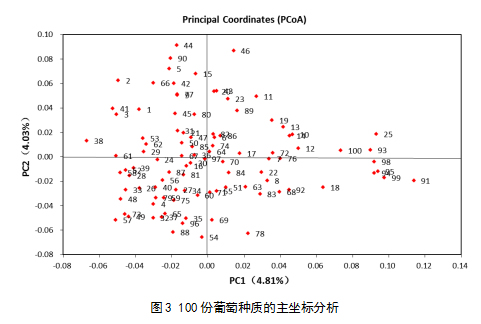
为了进一步了解不同葡萄种质间的的遗传关系,利用GenAlEx6.503基于遗传距离矩阵对100份葡萄种质资源进行了主坐标分析(PCoA)(图1)。结果显示变量和变量在两个主成分(PC)上的投影,PC1占总量的4.81%,表示两组间差异中可以解释全面分析结果的4.81%,PC2(主成分2)表示两组间差异中可以解释全面分析结果的4.03%,点与点之间的距离越小越相似,反之差异越大,从图中可看出两组间应该有明显的差异性。
2.3.3 遗传结构分析
利用STRUCTURE软件对100份葡萄种质进行遗传结构分析并预测其最佳K值,结果得出当ΔK达到最大值时(表5,ΔK=786.57),K的最佳分组数为2(图1A,1B),即葡萄核心集合主要分为两个集群。由于STRUCTURE软件分析支持主要群体之间差异,而一些群体之间的细微差异,尤其是那些混合群体之间差异并不明显。
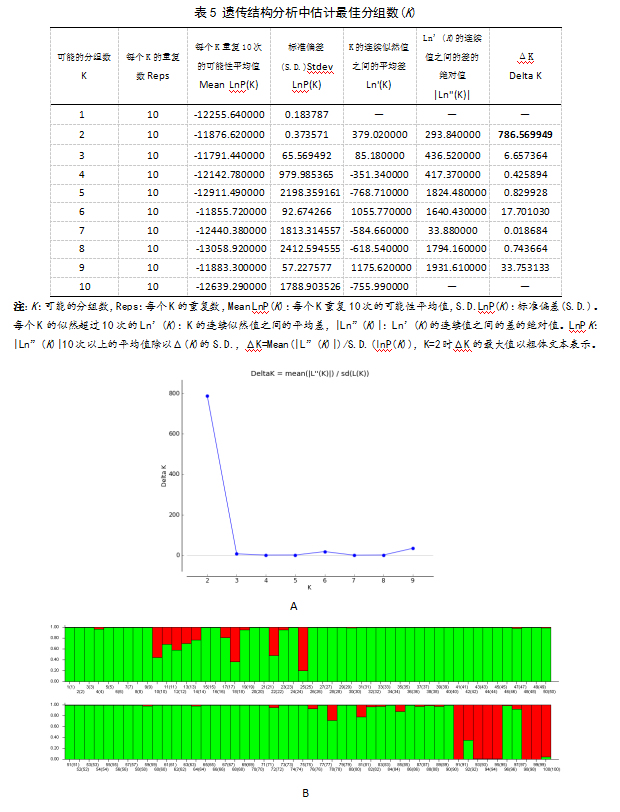

图4中不同的颜色块代表不同的群体,纵坐标表示各居群中的品种占某居群祖先成分的比例,横坐标为样品编号。从图4中可以看出根据各个品种的混合比率,将100份供试材料分成2个居群,图4B中,绿色板块代表欧亚种鲜食葡萄种群,红色板块代表欧亚种酿酒葡萄、欧美杂种和野生种质资源种群。
3 讨论
遗传多样性的研究对于了解物种的起源、适应性、基因资源分布情况、基因资源保护、维护物种稳定、挖掘优质资源、培育新种质等方面具有重要意义,通过对品种间亲缘关系的研究可以有效地进行亲本选配,并且对特殊种质进行保护。近年来SSR分子标记技术作为一种比较成熟的分子标记己广泛用于种质鉴定、亲缘关系分析、遗传多样性等方面的研究。本研究利用SSR毛细管电泳技术对100份葡萄种质资源亲缘关系和群体遗传结构进行了分析。100个样本中的33个SSR标记共检测到423个等位基因,每对引物扩增条带数4~26条,平均每对引物扩增条带数为12.82条。33对SSR引物在100份葡萄样品中扩增的等位基因组成中稀有等位基因数(≦1%)范围在0-9个,平均值为3.39,共有等位基因(1~≦20%)为8.26,最常见的等位基因数(>20%)为1.67。说明所选葡萄种质材料的遗传基础相对狭窄。本研究从70对SSR引物中筛选出33对能扩增出清晰稳定条带的引物,结果表明33对引物均能扩增出351条稳定清晰的DNA条带,多态性条带比率(PPB)为99.39%,每对引物扩增条带数4~21条,平均每对引物扩增条带数为10.64条,扩增条带大小范围为87~406bp。所有引物中,编号VMC9a2.1的引物扩增条带数最多,为21条,在引物评价种质资源遗传多样性的方法中,扩增条带数越多说明引物鉴别力越强,因此,VMC9a2.1为鉴别能力较强的引物,可作为种群内多样性鉴别的首选引物。检测到有效等位基因数(Ne)变化范围是1.582-12.305,平均为5.091;位点多态性信息含量(PIC);根据BOTSTEIN等提出衡量基因变异程度高低的多态性信息量指标PIC,本研究中PIC变幅为0.358-0.915之间,平均为0.716,均为高度或重度多态性信息引物。
葡萄种群的划分主要以其主要按原产地、形态解剖结构和亲缘关系为主要依据。本研究对100份葡萄种质资源的遗传多样性和群体结构进行了分析。进行了聚类分析,从聚类图2可以看出:(1)同一种类能够聚到一起,说明聚类结果可以很好的体现种质的种类或亲缘关系,但鲜食,酿酒,砧木等用途不能成为区别的有效依据。(2)不同地域的种质资源,但用途相同的也能够聚到一起,说明聚类结果不能很好的体现地域特性。例如,来源不同的酿酒品种聚在一起。(3)欧美杂种聚在一起,说明这些材料有相同或相近的亲本。(4)欧美杂种与欧亚酿酒葡萄品种及野生种质资源亲缘关系较近。分析原因可能有由于人工干预较少,含有古老的基因更相近的原因,但还有待进一步证明。在遗传水平上的分类结果与各种群分类基本一致,将100份葡萄种质资源分为两大类群,两大类群中又可分为4个亚群。其中欧亚种中鲜食和酿酒的较近,然后是欧美杂种和野葡萄和砧木类,这与唐宇宏等、刘茜等、方连玉等利用分子标记对葡萄资源亲缘关系的研究结果一致;而欧亚、欧美杂种与中国野生葡萄资源、砧木资源相距较远,此研究结果与温景辉等、张萌等的研究结果一致;聚类结果虽然与各种群的地理起源基本一致,但是也存在在欧亚种中出现1份砧木品种和1份野生种或在欧亚种中出现欧美杂种的现象,分析原因由于在扩增过程中不是全基因组序列,因此存在有些品种刚好没有表现出来的现象,具体原因还需进一步分析。另外,2个集群其中第一个主要为葡萄亚种鲜食葡萄资源,第二集群主要包含了欧亚种酿酒资源、欧美杂种、美洲种、中国野生种4个亚种群。从结果来看欧亚种的品种由于不同的用途面出现在不同的集群中;而另外的几个种也聚在了这个集群中。分析原因可能由于进化程度或进化方式不同导致含有古老或传统基因量多少存在差异引起的。
4 结论
利用33对SSR引物在100份葡萄品种进行了分析,从遗传结构分析并预测其最佳K的最佳分组数为2,即葡萄核心集合主要分为两个集群。2个集群其中第一个主要为葡萄亚种鲜食葡萄品种,第二集群主要包含了欧亚种酿酒品种、欧美杂种、美洲种、中国野生种4个亚种群。不同进化程度或进化方式在葡萄遗传结构中发挥了较重要的作用,影响了不同产地间葡萄种质资源的结构组成。