南宁综合试验站
时晓芳 韦荣福 黄桂媛 张瑛 林玲 韩佳宇 曹雄军 周思泓 王博 白先进 郭荣荣
摘要:【目的】探究矮壮素处理后夏黑葡萄二季果成花的主要生物学过程和重要基因,为促进二季果成花提供理论依据。【方法】以矮壮素处理后夏黑葡萄二季果第5-6节位冬芽为试验材料,采用转录组测序技术筛选差异表达基因(DEGs),并对其进行分析。【结果】与对照相比,矮壮素处理后-3 DAF、4 DAF、11 DAF冬芽样品中差异基因数分别为395、557和97。其中-3 DAF、4 DAF、11 DAF冬芽样品中分别有52、21、39个DEGs上调表达,343、536、58个DEGs下调表达,共有879个DEGs得到注释,富集最显著的Pathway主要有苯丙素和类黄酮生物合成、激素信号转导等。【结论】通过转录组测序和qRT-PCR验证发现,矮壮素处理夏黑葡萄后,二季果成花关键节位冬芽内苯丙素生物合成途径中的POD47和PODN1,类黄酮生物合成途径中的STS6,生长素信号转导通路中的IAA26,赤霉素信号转导通路中的GARP6 和GA3OX1等基因表达量发生明显变化,说明矮壮素促进成花的功能可能是通过改变树体内激素的含量及比值、次级代谢产物的合成和代谢来实现的。
关键词:葡萄;二季果;成花;矮壮素;分子机制
葡萄是世界上栽培面积最为广泛,人们最喜爱和加工产品最为丰富的水果之一。一年两收栽培是通过打破葡萄的正常生理周期,将本应在第二年春季萌发的冬芽在当年夏季修剪并用破眠剂打破休眠,当年抽生新梢,开启第二个生育周期并获得第二季冬果,达到增产与延长葡萄供应期等目的的有效途径。广西地处亚热带气候区,冬季较短,夏季较长,年平均气温在16-23 ℃之间,≥10 ℃的有效年积温为5036.7-8017.4 ℃,全年无霜期多在300d以上,适合研究和推广葡萄一年两收栽培技术。但是由于受到广西上半年多雨、少日照等自然条件的限制,部分品种生长旺盛,二季果成花不稳定或成花很少,在一定程度上阻碍了葡萄一年两收栽培技术的推广和优良品种的引进。夏黑葡萄是一年两收栽培种植较广的早熟葡萄品种之一,但是其冬芽分化不良、成花不稳定严重影响两收栽培的稳产、丰产。林玲等通过切片观察发现,夏黑葡萄新梢第5-6节位冬芽分化集中,成花率较高,且从初花期到盛花期大多数冬芽进入始分化期(始原基形成关键期),坐果期冬芽进入花原始体发育期到花序第一穗轴发育期(始原基分化关键期)。
对葡萄成花诱导中的激素含量分析结果表明细胞分裂素(CK)和赤霉素(GAs)是影响葡萄冬芽形成的两种主要激素。CK会诱导离体培养的幼嫩卷须分枝,并长出花序样的组织。而且在施加到茎尖时,CK会促进新形成的始原基发育成花序而不是卷须。此外,高浓度的GA3会造成主芽坏死,从而使副芽代替主芽而间接影响成花,因此,新梢旺长时坏死芽多。前期研究结果表明,矮壮素(CCC)作为GAs的一种拮抗剂,用其对夏黑葡萄叶片进行喷施处理,可以在花前3d和花后4d时降低新梢5-6节位冬芽内ZR/GAS比例,在花后11d时提高ZR/GAs的比例,最终增加二季果的成花率。但CCC处理增加二季果成花率的分子机制尚不明确,因此笔者通过转录组测序技术对CCC处理后夏黑葡萄二季果冬芽分化关键时期的差异基因进行分析,以期从分子生物学的角度进一步深入研究葡萄成花调控的机制,为葡萄的产期调节,尤其是一年两收栽培模式下二季果的生产提供理论基础指导,更好的为葡萄产业服务。
1 材料与方法
1.1 材料
供试品种为2014年定植在广西壮族自治区农业科学院明阳双季葡萄示范园内的夏黑葡萄,“V”形架避雨栽培,整形方式为“一”字型,栽植密度2.5m×1.5m,南北行向,滴灌。CCC(纯度98%)购自北京索莱宝科技有限公司。RNA提取试剂盒为Spectrum TM Plant Total RNA Kit 50 Prep, Sigma。
1.2 方法
1.2.1 CCC处理和样品采集
CCC处理夏黑葡萄叶片2次,分别在花前10d和花前1d进行,对夏黑葡萄全株叶片上喷施0.5g/L CCC。对照植株喷施纯净水。在对照和CCC处理的植株上,当主梢第14片叶展平时摘除其顶端,所有副梢全部抹除。于第一次CCC处理后7,14和21 d进行采样,分别为一季果花前3 d,花后4和11 d。(-3 DAF和4 DAF是始原基形成关键期,11 DAF始原基分化关键期)进行采集夏黑葡萄用于二季果生产的第5、6节位的冬芽,每个时间点,从6株植株上采集60个冬芽作为一个样品,每个处理重复3次。所有冬芽采集后迅速在液氮中速冻,并保存在-80℃。所有样品用来进行RNA-Seq建库和测序分析。
1.2.2 葡萄RNA的提取
葡萄冬芽总RNA用Spectrum TM植物总RNA提取试剂盒进行提取,然后用不含RNA酶的DNA酶除去基因组DNA。
1.2.3 转录组文库构建及测序
转录组文库构建参照Zhong等的方法,利用dUTP法构建Strand specific RNA-Seq文库,并使用Illumina HiSeq 2000平台对所建文库进行转录组测序。
1.2.4 差异表达基因的鉴定和比较分析
首先,利用Top Hat对转录组测序最终获得的高质量序列与葡萄参考基因组进行比对,根据比对结果参照V0版本的基因模型计算每个基因的原始表达。其次根据比对得到的mapped reads数与基因长度及比对到所有基因上的总reads数计算FPKM(fragments per kilobase of transcript per million fragments mapped)值,分析差异基因的表达量。然后应用 DESeq 软件包来处理原始表达数据,获得两个时间点之间每个基因的原始P值,经false discovery rate(FDR)多重校正获得corrected P值(Benjamini and Hochberg 1995)。最后将corrected P值小于0.05,两个时间点之间的FPKM比值(ratio)≥2或者≤0.5的基因鉴定为上调或者下调的差异表达基因。
利用在线分析工具Calculate and draw custom Venn diagrams (http://bioinformatics.psb.ugent.be/webtools/Venn/)获得不同样品间差异表达基因的交集和差集。
1.2.5 差异表达基因Gene Ontology(GO)分析和富集分析
将差异表达基因提交到在线分析工具Plant Met Gen MAP和Blast2Go系统计算每个GO term的基因数,从而得到具有某个GO功能的基因列表及基因数目统计。然后应用超几何检验,找出与整个基因组背景相比,在差异表达基因中显著富集的GO条目。计算得到的p-value通过FDR校正之后,以Q-value≤0.05为阈值,满足此条件的GO term定义为在差异表达基因中显著富集的GO term 。通过GO功能显著性富集分析来确定差异表达基因行使的主要生物学功能。
1.2.6 差异表达基因KEGG Pathway分析和富集分析
在生物体内,不同基因相互协调行使其生物学功能,基于Pathway的分析有助于更进一步了解基因的生物学功能。KEGG是有关Pathway的主要公共数据库。Pathway显著性富集分析以KEGG Pathway为单位,应用超几何检验,找出与整个基因组背景相比,在差异表达基因中显著性富集的Pathway。经过多重检验校正后,选择Q value≤0.05的Pathway定义为在差异表达基因中显著富集的Pathway。
1.2.7 差异基因表达模式聚类分析
对不同处理同一个时间点之间的差异表达基因取并集,利用Genesis软件中的K-means方法对这些差异表达基因进行表达模式聚类分析,了解CCC处理对夏黑葡萄冬芽分化过程中基因表达水平的影响。
1.3 差异基因实时荧光定量PCR验证
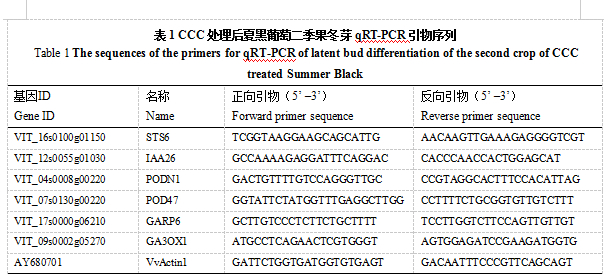
用PrimeScriptTM RTase试剂盒(TaKaRa, 大连)反转录合成cDNA,qRT-PCR使用LightCycler®480 SYBR® Green 1 Master 试剂盒(Roche, 瑞士),采用20μL反应体系:cDNA 模板1μL,2×SYBR Green Mix 10μL,上下游引物(10μmol·L-1)各1μL,超纯水7μL。 qRT-PCR 在LightCycler®480ǁ Real-Time PCR 仪(Roche,瑞士)上进行,反应程序为:95 ℃ 预变性1 min,95 ℃ 10 s,55 ℃ 30 s,72 ℃ 30 s,40个循环。利用Primer 6软件设计qRT-PCR引物,设计好的引物(表1)由奥科公司合成。内参基因为葡萄Actin1。基因的表达量采用相对定量的方法,即2-ΔΔCT 法。
2 结果与分析
2.1 CCC处理后夏黑葡萄二季果冬芽分化不同时期的转录组测序结果统计
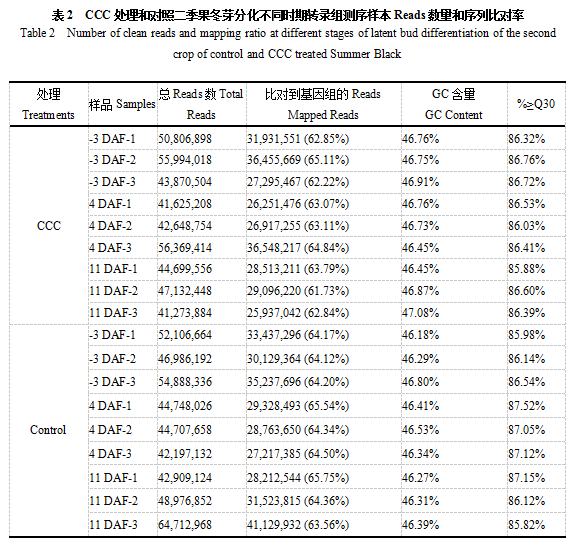
转录组测序结果显示,对照在三个时期分别获得153,981,192,131,652,816和156,598,944总Reads数,CCC处理在三个时期分别获得150,671,420,140,643,376和133,105,888总Reads数,这些高质量的reads中匹配到黑比诺葡萄基因组的匹配率在61.73%-65.75%之间(表1)。CCC处理和对照三个时期共18个样本的GC含量在46.18%-47.08%之间,较为一致(表1)。18个样本Q30百分比(评价质量值大于等于30的cycle所占的比例)≥85.82%,测序数据质量较高,可以满足后续分析的要求。
2.2 CCC处理后夏黑葡萄二季果冬芽分化关键时期差异表达基因筛选
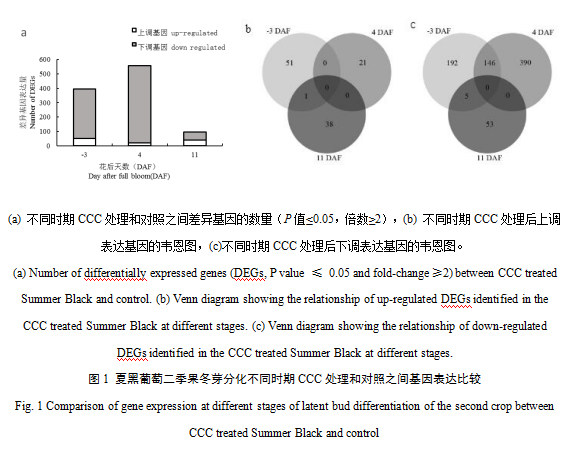
在-3 DAF,4 DAF和11 DAF,CCC处理和对照之间差异表达的基因数目分别为395,557和97。在三个时期,CCC处理后上调表达的基因数分别是52、21、39,下调表达的基因数分别是343、536、58,下调表达的基因数目均比上调表达的基因数目多,且CCC处理对冬芽内基因表达水平的影响在4 DAF最强,然后逐渐减弱(图1a)。同一时期CCC处理和对照之间的差异基因重叠情况如图1b, c所示。
2.3 CCC处理后夏黑葡萄二季果冬芽分化关键时期差异基因的表达模式聚类分析
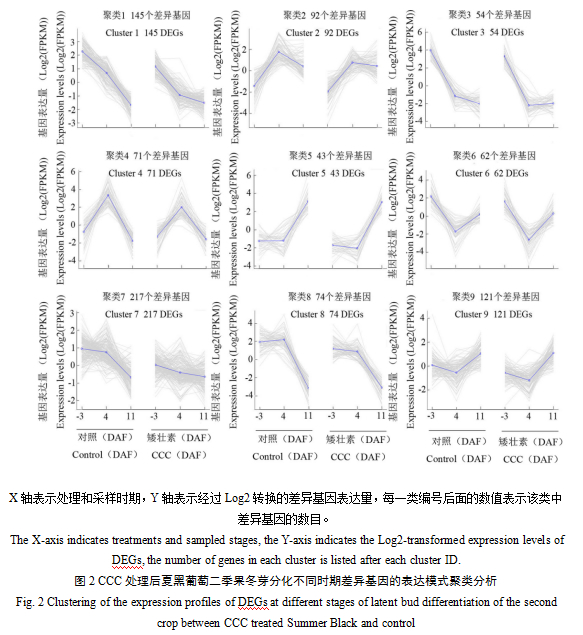
从图2可以看出,对CCC处理和对照之间不同时期的所有差异基因取并集,共获得879个差异基因, cluster 1、5、6、7和8中的基因在-3 DAF和4 DAF下调表达,cluster 2、3、4和9中的基因在4 DAF下调表达。CCC处理后差异基因的表达趋势与对照相似,大多数差异基因的表达量比对照较低。
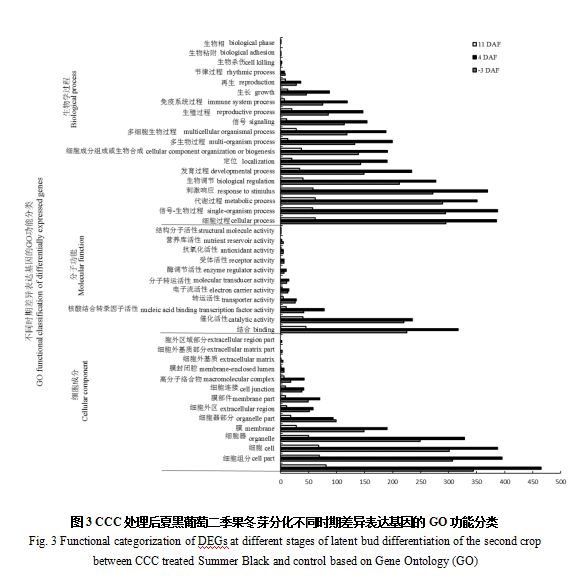
2.4 CCC处理后夏黑葡萄二季果冬芽分化不同时期差异表达基因的GO功能分类
对CCC处理后夏黑葡萄二季果冬芽分化不同时期与对照之间的差异表达基因进行了GO生物过程功能分类,并将其分为生物学过程,分子功能和细胞成分三类(图3)。结果表明,在CCC处理后夏黑葡萄二季果冬芽分化的不同时期,参与细胞组分的差异表达基因数目最多,参与细胞和细胞过程的差异表达基因数目次之,还有一些参与信号-生物过程,代谢过程,刺激响应,细胞器,结合,催化活性,生物调节,发育过程,定位,细胞成分组成或生物合成,多细胞生物过程,多生物过程,信号,膜等的差异基因。
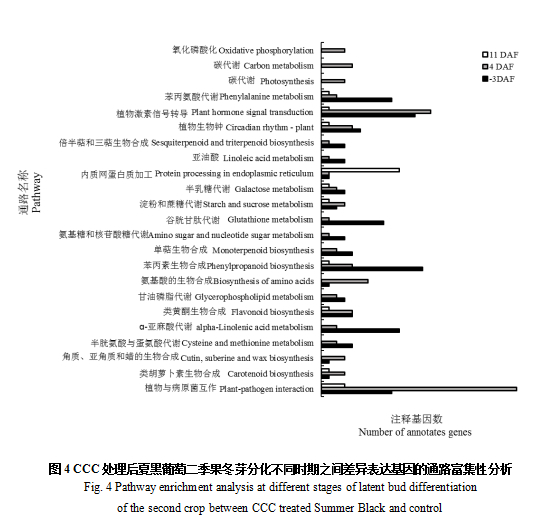
2.5 CCC处理后夏黑葡萄二季果冬芽分化不同时期差异表达基因的代谢通路富集性分析
对CCC处理后夏黑葡萄二季果冬芽分化不同时期与对照之间的差异表达基因进行了合成和代谢通路富集性分析。如图4所示,-3 DAF时,CCC处理与对照之间的差异表达基因显著富集的通路大多数与苯丙素生物合成、植物激素信号转导、ɑ-亚麻酸代谢、苯丙氨酸代谢、谷胱甘肽代谢、植物与病原菌互作以及植物生物钟相关。4 DAF时,CCC处理与对照之间的差异表达基因显著富集的合成和代谢通路大多数与植物-病原菌互作、植物激素信号转导、氨基酸合成、苯丙素生物合成、碳代谢、植物生物钟以及类黄酮生物合成有关。11 DAF时,CCC处理与对照之间的差异表达基因显著富集的合成和代谢通路大多数与内质网中的蛋白质加工和植物-病原菌互作有关。其中,氧化磷酸化、碳代谢和光合作用只在4 DAF时在CCC处理与对照之间的差异表达基因中富集。
2.6 CCC处理后夏黑葡萄二季果冬芽分化重要响应基因的分析
根据上述对所有差异表达基因的GO功能分类和通路富集分析,发现可能参与CCC处理后冬芽分化的主要有花发育,正调控花发育及负调控花发育等GO功能分类,可能参与的主要通路有植物生物钟/植物激素信号转导/苯丙素合成/类黄酮生物合成等。本研究对这些功能分类和通路中重要响应的差异基因及差异表达的转录因子进行详细分析。
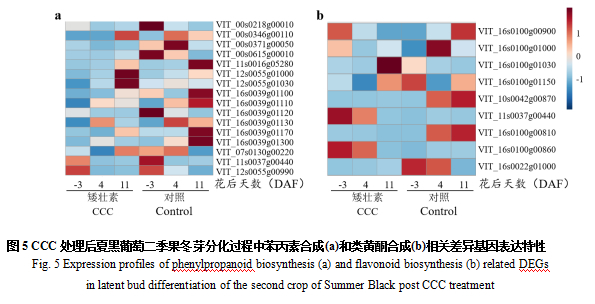
2.6.1 苯丙素合成和类黄酮合成相关差异基因筛选及表达模式分析
本研究发现CCC处理会影响夏黑葡萄二季果冬芽分化过程中相关基因的表达水平(图5)。如图5a所示,在-3 DAF时,CCC处理会降低苯丙素生物合成通路中过氧化物酶25和N1以及PAL的表达水平,此外,苯丙素生物合成通路中的甘露醇脱氢酶和类黄酮生物合成通路中的芪合成酶3,4,6的表达水平也下调;4 DAF时,苯丙素生物合成通路中PAL、过氧化物酶47和阳离子过氧化物酶2的表达下调,苯丙素生物合成通路和类黄酮生物合成通路中的莽草酸O-羟基肉桂转移酶,以及苯丙素生物合成通路中的甘露醇脱氢酶和类黄酮生物合成通路中的芪合成酶3,4,5的表达水平也下调;而在11 DAF时,苯丙氨酸代谢和苯丙素生物合成通路中的过氧化物酶N1和47,阳离子过氧化物酶2,以及类黄酮生物合成通路中的芪合成酶4和6的表达上调,苯丙素生物合成通路中的过氧化物酶25和PAL,以及类黄酮生物合成通路中的芪合成酶3、4和查尔酮合酶17的表达下调。
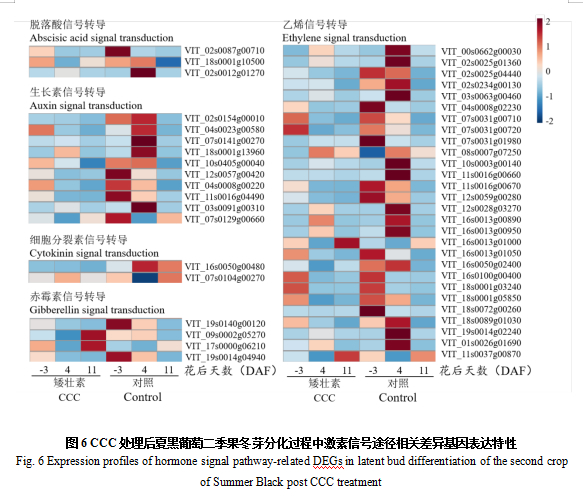
2.6.2 植物激素信号途径相关差异基因筛选及表达模式分析
本研究筛选了脱落酸、生长素、赤霉素、细胞分裂素和乙烯等5种可能涉及CCC处理促进葡萄二季果成花的调控过程,对CCC处理后激素信号途径相关基因的表达水平进行了分析。
研究结果表明,ABA信号途径中的ABA受体PYL1在-3 DAF上调;ABA 8'-羟化酶4在11 DAF上调。生长素信号途径中的生长素诱导蛋白X10A在-3 DAF上调,生长素诱导蛋白22D和生长素响应蛋白IAA26在11DAF上调。细胞分裂素信号途径中的细胞分裂素羟化酶在-3 DAF下调,在11 DAF上调。赤霉素信号途径中的赤霉素调节蛋白6在-3 DAF上调,赤霉素3-β-双加氧酶1和赤霉素调节蛋白6在11 DAF上调。乙烯信号途径中DEGs数目较多,其中,RAP2-11和ERF5在-3 DAF上调,ABR1在-3 DAF下调;ERF017、ERF109、ERF016、ERF022、ABR1和一个AP2类的乙烯响应转录因子在4 DAF显著下调;ERF2、ERF5、ERF109和ABR1在11 DAF上调(图6)。
2.6.3 成花相关差异基因筛选及表达模式分析
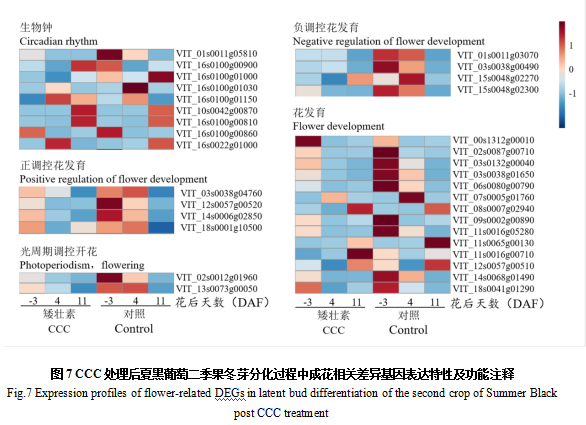
为进一步明确夏黑葡萄二季果的成花调控机理,本研究对CCC处理后冬芽分化关键时期成花相关基因的表达水平进行了分析。结果显示,CCC处理后,在-3 DAF时花发育相关基因bHLH39的表达水平上调。在4 DAF时,生物钟途径中的查尔酮合酶17上调表达,生物钟途径中的芪合成酶4和5、花发育相关基因bHLH39下调表达。在11 DAF时,生物钟途径中的芪合成酶4和6,花发育相关基因细胞色素P450,正调控花发育的bHLH63、BEE1和ABA 8'-羟化酶4,以及负调控花发育的TEM1上调表达(图7)。
2.6.4 差异表达转录因子筛选及表达模式分析
转录因子在植物的生长发育中起重要作用。本研究通过分析夏黑葡萄二季果冬芽分化关键时期CCC处理后差异表达的转录因子,发现有23个转录因子家族的116个基因响应CCC处理。其中,编码差异基因数目最多的转录因子为AP2/ERF-ERF家族成员,其次是WRKY、NAC、bHLH、MYB和Tify家族成员。在-3 DAF时,1个C3H家族成员受CCC诱导显著上调表达;此外,有2个AP2/ERF-ERF基因,1个bHLH基因,1个HSF基因,1个LOB基因,1个Tify基因,1个WRKY显著下调表达。在4 DAF时,只有Trihelix家族的一个差异基因在CCC处理后上调表达,下调表达倍数小于1/8的DEGs包含1个AP2/ERF-AP2基因、4个AP2/ERF-ERF基因、1个C2H2、1个NAC和1个Tify。在11 DAF时,1个bZIP家族成员在CCC处理后显著上调表达(图8)。

2.6.5 CCC处理后夏黑葡萄二季果冬芽差异基因qRT-PCR验证转录组结果
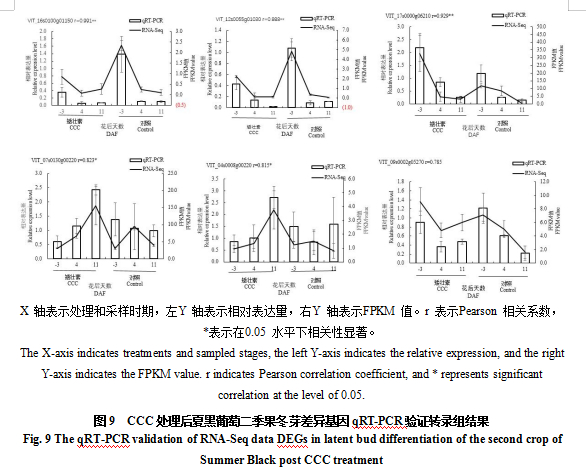
从代谢通路中筛选出6个相关的候选基因POD47、PODN1、STS6、IAA26、GARP6、GA3OX1,通过qRT-PCR分析在CCC处理后不同采样时期的表达水平,并与转录组结果进行比较。结果如图9所示,Pearson相关分析显示,有5个候选基因的qRT-PCR与转录组测序结果的相关性大于0.8,1个候选基因的qRT-PCR与转录组测序结果的相关性为0.785,基因表达趋势基本一致,说明转录组FPKM值相对准确。
3 讨 论
近年来,葡萄一年两收栽培技术在我国南方地区得到了大面积的推广和应用,但是由于南方大部分地区春季阴雨天气较多,易造成冬芽分化不好的问题,最终影响二季果的成花和产量。前人研究表明CCC处理会促进葡萄花序诱导。本研究以CCC处理后的夏黑葡萄二季果生产关键节位的冬芽为试材,通过Illumina HiSeq 2000高通量测序技术来揭示CCC处理促进葡萄成花的机制。
本研究发现在夏黑葡萄二季果冬芽分化关键时期,CCC处理会诱导苯丙素合成和类黄酮合成通路中部分基因的上调或下调表达,表明这2个通路可能参与调控葡萄冬芽分化,但其具体的调控机制还需要进一步更深入的研究。
前人研究发现植物激素在木本植物芽生长发育、成花诱导及树体生长发育中有重要作用。拟南芥内源GA合成受阻或破坏GA在体内的信号转导过程,开花进程均会受到影响;GA对苹果、梨、红灯樱桃等多年生果树的花芽分化有抑制作用,而CTK则促进其花芽分化;高水平ZT/GA3、IAA/ABA比值有利于蒿柳花芽的形态分化。Xing 等通过分析苹果花芽形成不同时期的转录组数据,发现CK、ABA和GA信号转导途径组成的调控网络参与苹果的成花诱导。筛选响应CCC处理与植物激素信号途径相关的差异表达基因,可以为进一步解释植物激素在参与CCC促进夏黑葡萄二季果成花调控的分子机理提供依据。
葡萄花序的形成主要在两个时期受到调节:始原基的形成时期和始原基的分化时期,其中GA和CK是成花的关键调节剂。CK会促进始原基分化为花序而不是卷须。前期研究结果表明在-3 DAF和4 DAF时,CCC处理会通过降低ZR含量及ZR/GAs的比值,促进始原基形成,而在11 DAF时,CCC处理会使ZR/GAs的比值增高,从而促进始原基分化为花序。本研究发现CCC处理后,在-3 DAF时,赤霉素调节蛋白6的表达上调,细胞分裂素羟化酶的表达下调;而在11 DAF时,赤霉素3-β-双加氧酶1、赤霉素调节蛋白6和细胞分裂素羟化酶均上调表达(图6),进一步表明CCC处理会通过调控GA和CK信号途径中相关基因的表达水平,从而改变这两种激素的含量,进而调控始原基的形成和分化方向。
芽内IAA含量较高会刺激芽的生长并促进成花诱导的起始。与成花较差的‘长富2号’相比,易成花的‘秦冠’苹果芽内IAA生物合成的关键基因的表达上调。本研究发现CCC处理后夏黑葡萄二季果成花关键节位芽内IAA诱导蛋白X10A和22D,以及IAA响应蛋白IAA26的表达有所上调(图6),表明IAA在调控夏黑葡萄二季果冬芽成花的过程中发挥作用。
作为TCA循环的下游信号通路,ABA信号转导通路在调控成花诱导和花形成中的主要功能包括调控植物生物钟及光周期相关基因的表达。本研究发现CCC处理后在-3 DAF时ABA受体PYL1的表达上调;4 DAF时生物钟途径中的查尔酮合酶17上调表达;11 DAF时ABA 8'-羟化酶4以及生物钟途径中的芪合成酶4和6的表达上调(图6-7)。此外,ABA相关的糖信号还会影响植物的营养生长、开花时间及胁迫响应,表明ABA在调控植物成花诱导的过程中是多个信号通路(如:糖、光周期和生物钟)的中心因子。
CCC处理后,除了生物钟途径中的部分基因在4 DAF或11 DAF上调表达外,在-3 DAF时花发育相关基因bHLH39的表达水平轻微上调;11 DAF时,花发育相关基因细胞色素P450,正调控花发育的bHLH63、BEE1和ABA 8'-羟化酶4,以及负调控花发育的TEM1均上调表达(图7)。表明CCC处理可能会通过上调或下调成花相关基因的表达来调控始原基的形成和分化。
转录因子在植物生长发育中发挥重要作用。Tsai等构建了小兰屿蝴蝶兰成熟花芽的cDNA文库,通过分析鉴定出来的ESTs序列,发现C3H和trihelix基因家族占鉴定出的转录因子的25%。通过分析腊梅中的两个C3H类锌指转录因子CzCPF1和CzCPF2在花发育中的功能,发现这两个基因的表达模式与花原基的分化有关。本研究通过分析CCC处理后夏黑葡萄二季果冬芽分化关键时期差异表达的转录因子的表达模式,共鉴定到14个转录因子家族的89个差异表达的转录因子(图8)。部分差异表达的转录因子,如bZIPs、ERFs、MYBs等参与激素响应,WRKYs和NACs则主要参与胁迫响应。还有一些转录因子在多个开花途径中直接调控开花,如C2H2家族的IDD转录因子参与蔗糖途径中的开花调控,NF-YB调控光信号途径中的开花。因此,CCC处理后夏黑葡萄二季果成花关键节位芽内差异表达的转录因子可能参与多个生物学过程中的成花调控。
4 结 论
本研究通过对CCC处理后夏黑葡萄二季果冬芽分化关键时期(始原基形成关键期和始原基分化关键期)18个样本的RNA-Seq数据分析和qRT-PCR验证发现,响应CCC处理的差异表达基因,主要涉及苯丙素生物合成(POD47和PODN1)、类黄酮生物合成(STS6),激素信号途径(IAA26、GARP6和GA3OX1)及差异表达的重要转录因子,表明CCC处理作为一种化学措施,其促进成花的功能可能是会通过改变树体内激素的含量及比值,次级代谢产物的合成和代谢,以及成花相关基因的表达模式。正是这些响应CCC处理的差异表达基因形成系统的、相互联系的复杂基因调控网络共同促进葡萄二季果的成花。