酿酒微生物岗位
1 引言
葡萄酒是包含多种无机和有机成分的复杂基质,这些化合物来自葡萄、发酵和陈酿。即一类香气是从葡萄中提取的香气,二类香气是葡萄酿造过程中由酵母或细菌代谢的香气,三类香气是由橡木提取或是葡萄酒储存过程中的化学反应产生的香气。在葡萄酒中发现了几种类型的挥发性化合物:醇类、脂肪族、苯衍生物、羰基、酯类、有机酸、含硫化合物、莽草酸衍生物、多元醇、异戊二烯衍生萜烯和C13-降异戊二烯、甲氧基吡嗪、乙烯基和挥发性酚类(Bisson and Karpel2010)。源自发酵的挥发性化合物占葡萄酒总香气成分的最大百分比,酵母发酵产生的香味活性物质分为六大类:有机酸、高级醇、羰基化合物、含硫分子、酚类化合物和挥发性酯。陈酿是某些葡萄酒最终风味特性的关键步骤。在橡木桶中老化过程中,木桶将一系列芳香物质转移到葡萄酒中。从木桶中提取的挥发性化合物主要是糠醛化合物,例如愈创木酚,橡木或威士忌内酯,香兰素或丁香醛。除此之外,一些微生物衍生的化合物,如多糖,游离氨基酸和肽,也会释放在葡萄酒中。这些化合物来自死亡的酵母和细菌(酒泥),在裂解过程中还可能释放一些活性酶不断改变酒的风味(Ferreira & Cacho2009)。挥发性化合物可以直接合成或提取,也可以以非挥发性前体的形式存在,需要化学或生化活性来释放气味活性成分。
2 品种特征香气
葡萄衍生的成分是葡萄酒的主要香气或品种特征的主要来源,主要分布在果皮组织中。这些化合物大多以糖或氨基酸结合物的形式存在于外果皮细胞的液泡中,有些则以自由挥发物的形式存在。以偶联物形式储存在外果皮细胞液泡中的化合物通过糖苷酶和肽酶的作用被释放,糖苷酶和肽酶是在压榨和发酵过程中引入的,从而增加了成品酒中可感知的量。与葡萄品种香气有关的挥发性化合物包括萜烯类、降异戊二烯和吡嗪类(Villamor andRoss 2013)。特定的芳香化合物或前体通常存在于几种葡萄品种的葡萄酒中。每个葡萄品种独特的芳香个性是由于它的各种化合物的特殊组合。葡萄果实似乎缺乏解剖结构来储存亲脂性挥发性有机化合物,这些化合物往往以水溶性糖苷或与半胱氨酸等氨基酸结合的形式储存。糖苷酶和从这些水溶性形式中释放挥发性香气化合物的肽酶在葡萄酒的风味和香气中起着至关重要的作用。
单萜类在很大程度上定义了与马斯喀特有关的葡萄酒,而果味的挥发性硫醇则定义了与长相思有关的葡萄酒(SWIEGERS et al. 2005)。甲氧吡嗪(MPs)是长相思白葡萄酒和葡萄酒中“生青”味的主要来源(吕佳恒 et al. 2017)。在以赤霞珠葡萄(主要产于葡萄皮)中制成的赤霞珠葡萄酿造的葡萄酒中,已鉴定出高度调味的化合物2-甲氧基-3-异丁嗪。尽管葡萄酒的阈值更高,但它赋予了各种各样描述的“青草”或“甜椒”香气。甲氧吡嗪是含氮的环结构,是葡萄中氨基酸分解代谢的副产物。化合物2-甲氧基-3-异丁基吡嗪(IBMP)是长相思白葡萄酒的草味,青椒,辣椒和芦笋香气的主要来源。在长相思葡萄酒以及葡萄酒中,定量发现IBMP是最高浓度的吡嗪,其他浓度较低的吡嗪为2-甲氧基-3-异丙基吡嗪(IPMP),浓度较小的2-甲氧基-3-仲丁基吡嗪(SBMP)。许多研究人员已经在葡萄和葡萄酒中报道了脂肪族C6化合物,在有空气的情况下通过不饱和脂质的酶促氧化。在压榨葡萄的过程中,C6醛的形成非常迅速。C6醛是葡萄浸软葡萄叶中最丰富的挥发物,被迅速转化为己醇,此过程在存在酵母的情况下得以加速(Hashizume andSamuta 1997)。
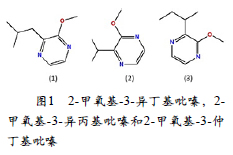
麝香味的品种有最高浓度的萜烯糖苷(Terpene glycosides)。在发酵过程中,通过酵母溶菌酶和酸性发酵条件(pH ~3.5)水解戊糖苷释放挥发性萜烯(González-Barreiro etal. 2015),最重要的是芳樟醇,香叶醇,橙花醇和香茅醇。
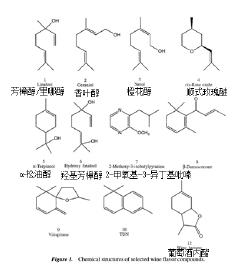
木桶中的陈酿是用来稳定颜色,改善葡萄酒的清爽性和感官特性的过程。酒的香气成分也可能会受到橡木桶中存储的影响,因为酒桶为葡萄酒提供了令人赏心悦目的香气,例如椰子,香草,
丁香,烟熏和木材。在陈酿过程中,许多化合物从木材转移到葡萄酒中:多酚,内酯,香豆素,多糖,烃和脂肪酸,萜烯,C13-去甲肾上腺素,类固醇,类胡萝卜素和呋喃化合物。木材挥发物,例如顺式和反式 -β-甲基-γ-八内酯(橡木内酯),丁香醛,香兰素,松柏树醛,芥子醛及其醇类,丙丁香酮和丙香烷酮,羟基-大麦草酮-2-酮和羟基-大麦草酮-3-酮,糠醛,5-甲基糠醛,愈创木酚,丁子香酚,4-乙基苯酚和4-乙基愈创木酚,糠醇,β-紫罗兰酮,γ-壬酸内酯和乙酰香草醛赋予典型陈年葡萄酒的感官特性。陈酿中,最重要的化合物之一是β-methyl-γ-octalactone,称为橡木内酯(oaklactone)或 威士忌内酯(whiskeylactone)。橡木内酯有两种同分异构体,这两种异构体都有一种木质、橡木、椰子般的香气。然而,顺式异构体的香气阈值为92 ppb,而反式异构体的香气阈值为460ppb。橡木内酯顺式和反式的比例可以用来区分葡萄酒在美国还是法国/欧洲橡木桶中发酵。木材还可以吸附某些香气化合物(如2-苯基醇,癸酸乙酯),从而改变其在溶液中的浓度。
TDN为陈酿的雷司令贡献了一种典型的柴油或类似煤油的芳香,并具有阈值20 ppb。4-巯基-4-甲基戊烷-2- 1 - 13 (MMP)为Scheurebe葡萄酒增添了典型的黑醋栗味。
3 主要发酵香气
葡萄酒发酵过程中,糖酵解途径和代谢途径在维持酵母的生长外,产生了改变葡萄酒风味的挥发性和非挥发性代谢物。影响这些化合物的因素包括:营养条件、发酵条件和发酵菌种。
3.1 羰基化合物(carbonyl)
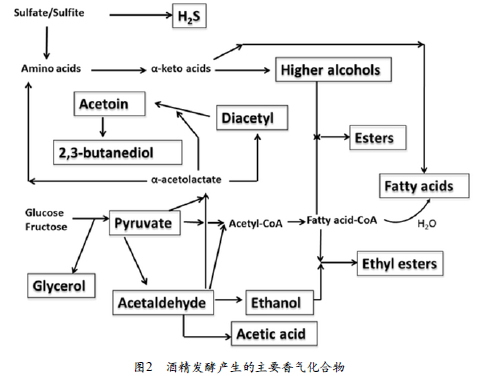
从数量上看,糖酵解的直接产物和副产物代谢产物的浓度最高。这些化合物包括乙醇,甘油和乙酸。发酵和储存过程中(醋酸杆菌)的氧化反应(Polášková et al.2008)会导致乙醛(Acetaldehyde)的形成,从而产生许多陈年葡萄酒和雪利酒典型的坚果味和雪利酒味。与氧化有关的香气性质的变化可能与醛的形成有关。醛类化合物对葡萄酒的香气很重要,因为它们的阈值很低。乙醛也是葡萄酒酿造过程中由丙酮酸盐(图2)形成的重要香气化合物,占葡萄酒总醛含量的90%以上。含量低时,该化合物可为葡萄酒和其他饮料带来令人愉悦的水果香气,但含量高时,则变成刺鼻的刺激性气味(Styger et al.2011)。由乙醛形成的重要气味物质是二乙酰(Diacetyl)。二乙酰基主要由苹果酸乳酸发酵过程中的乳酸菌形成,但酵母也能够在酒精发酵过程中合成该化合物。大多数的二乙酰基被进一步代谢为丙酮酸和2,3-丁二醇。二乙酰基在低浓度带给葡萄酒酵母,坚果,烤面包味,但在高浓度下,它具有与乳酸相关联的黄油香气。
3.2 高级醇
葡萄酒发酵伴随着脂肪醇和芳香醇的形成,称为杂醇或高级醇。主要的脂肪醇包括丙醇,异戊醇,异丁醇和活性戊醇,主要的芳香醇为2-苯乙醇和酪醇。葡萄酒中主要的高级醇是异丁醇,苯乙醇和异戊醇(Belda et al. 2017)。脂肪醇是通过Ehrlich途径从氨基酸分解代谢中衍生的,主要包括三个步骤:转氨、脱羧和还原(由BAT2,PDC1和ADH1基因编码)。尽管对这种途径的研究和讨论最多,但在氨基酸的上游(合成代谢途径)生物合成过程中也会形成高级醇。最重要的是通过异亮氨酸-亮氨酸-缬氨酸(ILV)途径从头开始合成支链氨基酸(BCAA)。
Ehrlich途径的第一步涉及由基因BAT1(TWT1或ECA39),BAT2(TWT2或ECA40),ARO8和ARO9基因编码的四种酶。这些酶是转氨酶,使用谷氨酸/α-酮戊二酸酯作为供体/受体,催化胺在氨基酸及其各自的α-酮酸之间的转移。转氨后,剩余的α-酮酸可以脱羧形成相应的醛,这是Ehrlich途径中的不可逆反应。酿酒酵母中有五个编码脱羧酶的基因:三个编码丙酮酸脱羧酶的基因(PDC1,PDC5和PDC6),ARO10和THI3。从这些基因中,仅PDC5和ARO10被描述为编码具有广泛底物特异性的脱羧酶。脱羧后,杂醇醛进入Ehrlich途径的最后一步,在此过程中,它们通过醇脱氢酶转化为各自的醇。由ADH1,ADH2,ADH3,ADH4和ADH5编码的酿酒酵母酒精脱氢酶中的任何一种或由Sfa1加密的甲醛脱氢酶均能够催化杂醛转化为高级醇(Pires et al. 2014)。
氨基酸分解代谢由NCR(氮分解代谢物阻抑)控制,NCR是一个复杂的调节系统,使酵母能够选择提供最佳生长的氮源。NCR主要由四个转录因子(GATA因子)以及调节蛋白Ure2p介导。
3.3 挥发性酚
异味通常被描述为动物、马厩、和马汗,可能是葡萄酒中挥发性酚类物质的贡献,这些物质的含量高于阈值水平。乙烯基酚和乙基酚在这类化合物中是重要的,挥发性酚4-乙基苯酚(4-EP),4-乙基愈创木酚(4-EG),4-乙烯基苯酚和4-乙烯基愈创木酚源自木材老化。这些化合物描述为“动物”和“酚”的感官特性。它们是由酿酒厂中的污染物,如酒香酵母(Brettanomyces)和德克酵母(Dekkera),通过脱羧和还原葡萄中存在的阿魏酸(ferulic)和对-香豆酸(p-coumaric acids)而产生的。
葡萄酒中4-EP和4-EG的感官阈值分别为440μg/ L和33μg/ L。4-EP和4-EG以不同的数量和比例出现在葡萄酒中,具体取决于所使用的葡萄品种和葡萄酒风格。平均而言,它们分别以10:1的比例出现,这对应于对香豆酸和阿魏酸的相应前体比例。另一项研究表明4-EP与4-EG的平均比例随葡萄酒类型的不同而不同,赤霞珠赤霞珠约为10:1,西拉子约为9:1,梅洛为8:1,黑比诺为3.5:1(Wedral etal. 2010)。
任何风味化合物在高浓度时都可能是不受欢迎的。呋喃酚furaneol (2,5-二甲基-4-羟基-2,3-二氢-3-呋喃酮,2,5-dimethyl-4-hydroxy-2,3-dihydro-3-furanone),是一种不受欢迎的草莓味。一些酿酒品种和美洲种(V. vinifera-V.labrusca)杂交种保留了这种呋喃酚异味。
3.4 乙酸酯
酯可分为两类:酶促形成的和葡萄酒陈酿过程中形成的。酶促酯的合成是由许多食物微生物产生的酯酶和脂肪酶催化的。酶形成的酯的实例包括乙酸乙酯,丁酸乙酯,己酸乙酯和辛酸乙酯。由于其复杂的性质,葡萄酒在储存期间甚至在开瓶后也会不断发生成分变化。这可能是由于水解和酯化作用,也可能是由于与羟基自由基相关的过程将酯氧化而引起的。乙酸酯由酸基(乙酸)和醇基组成,该醇基是乙醇或氨基酸代谢衍生的复合醇,包括乙酸乙酯、乙酸异戊酯、乙酸异丁酯、乙酸己酯等(Sumby et al.2010)。
3.5 乙基酯
酯类物质在酒精发酵,苹果酸发酵和陈酿过程中形成。乙酰CoA羧化酶将乙酰CoA转化为丙二酰CoA,该过程是一个限速步骤(Li et al. 2014)。酿酒酵母乙酰CoA羧化酶(ACC)具有四聚体结构,每个亚基有一摩尔共价结合的d-生物素(Wakil et al. 1983)。
随后丙二酰辅酶A脱羧与KS发生缩合反应。KR、DH和ER亚基串联催化形成饱和的酰基-载体蛋白(ACP),成为进一步添加脱羧丙二酰-ACP的底物。乙酰辅酶A和丙二酰辅酶A作为前体物质被不断吸收参与酰基-ACP的延长(Ferreira etal. 2018)。其中,酰基-ACP携带从一个酶活性位点到另一个酶活性位点的脂肪酰基中间体。在脂肪酸骨架上每延伸双碳单元即消耗两摩尔NADPH,并释放CO2(Radakovits etal. 2010)。在酿酒酵母中,NADPH主要由胞质溶胶中的戊糖磷酸途径产生。脂肪酸的合成取决于高能量负荷条件。在脂肪酸合成中,酮合酶,硫酯酶,丙二酰-棕榈酰转移酶(MPT)和乙酰转移酶(AT)决定脂肪酰链长度。FAS和ATP-CL的多酶复合物受到存在于培养基中的外源长脂肪链(例如FAs,正链烷等)的抑制(Meyer and Schweizer1976)。
葡萄酒中酯类的净积累量取决于酵母中酯合成酶和酯酶之间的平衡。由于挥发性酯随着时间的推移会逐渐水解,因此有时认为挥发性酯对葡萄酒的有益效果并不重要。实际上,取决于酸-酯平衡,支链脂肪酸乙酯会在葡萄酒陈酿过程中增加。葡萄酒中所发现的酯水平是由合成、酶促或自发水解以及底物分子水平决定的。在一些陈年葡萄酒中,酯类化合物由于完全水解而对香气没有贡献,而在另一些陈年葡萄酒中,酯类化合物对葡萄酒风味有重要贡献已鉴定出与酯形成有关的基因。乙酯的形成是由EEB1、EHT1和YMR210W编码的酰基转移酶催化的,而乙酸酯是由醇转移酶ATFI和ATF2的作用下形成的。这些基因的突变分析表明,其他未知的转移酶也可能参与了酯的合成。酵母菌也具有酯水解酶。主水解酶由IAHll基因编码。EEBI和EHT1也具有酯酶活性。生物合成或杂醇油的形成也会通过消除核心反应物而对酯的形成产生负面影响(Saerens et al. 2009)。肉碱乙酰转移酶的过表达减少了酯形成,提示酯生物合成可能被破坏。
酵母中有三种肉碱乙酰转移酶:CAT2、YAT1和YAT2。温度变化对酯的形成有显著影响。温度越低,酯含量越高(Saerens et al. 2009)。研究表明,这种效应不仅仅是由于挥发性酯的保留或酯酶活性的抑制,而是由于基因在低温条件下的表达模式(Beltran et al. 2006)。
3.6 含硫化合物(S-containingvolatile)
与诸如甲氧基吡嗪之类的香气化合物不同,葡萄汁中几乎不存在挥发性硫醇(Thiols)。葡萄酒中含硫化合物的存在有两个主要来源:(1)非酶促过程,包括酿造和储存过程中含硫化合物的化学反应(例如光化学反应);(2)酶促过程,包括酵母和乳酸菌发酵过程中含硫氨基酸的降解。
葡萄酒中的挥发性硫化物可分为两类。一方面,某些挥发性的硫化合物可能会产生负面的香气,例如腐烂的鸡蛋,这是由葡萄酒酵母形成H2S引起的。H2S的产生涉及几个因素:元素硫的高残留水平,二氧化硫的存在,有机硫化合物的存在,高浓度的氨基酸如苏氨酸,蛋氨酸或半胱氨酸以及氮的限制。硫的另一种来源是它对微生物生长的抵抗,例如焦亚硫酸钾。除了这些来源,主要的H2S产生发生在从无机或有机硫源合成含硫氨基酸蛋氨酸和半胱氨酸的过程中。在酿酒酵母中,H2S是通过硫酸盐还原序列(SRS)途径产生的。当存在对半胱氨酸和蛋氨酸代谢的需求,其通常在SRS途径被诱导。整个硫代谢途径也通过半胱氨酸细胞内浓度控制的几种机制进行调节。这些机制涉及硫酸同化和含硫氨基酸合成基因的转录,这些基因主要受MET4和GCN4基因调控。
另一方面,有称为“热带挥发性硫醇”的化合物:4-巯基-4-甲基戊烷-2-酮(4-MMP),3-巯基己醇(3-MH)和3-巯基乙酸乙酯(3-MHA)。白葡萄酒品种香气的其他相关硫醇为4-巯基-4-甲基戊-2-醇(4-MMPOH)和3-巯基-3-甲基丁-1-醇(3- MMB)。在酒精发酵过程中,芳香族硫醇通过酵母活性从其非芳香族前体中释放出来。结构上与半胱氨酸相似的Cys-3MH前体的摄取是由氨基酸转运蛋白诱导的。半胱氨酸化的前体一旦进入细胞,就会被碳硫β-裂合酶裂解生成有气味的挥发性硫醇。被鉴定为β-裂合酶的四个基因(BNA3,CYS3,GLO1和IRC7)影响挥发性硫醇4MMP的释放。大多数菌株酿酒酵母中呈现38-bp的IRC7序列碱基对缺失,其编码功能更少的酶。STR3β-裂合酶也负责硫醇的释放,但比活性较低。3MHA通过3MH与乙酸的酯化反应形成。该反应由形成酵母酯的醇乙酰基转移酶控制,该酶由ATF1基因编码。
3.7 萜类物质(terpenes)
萜类化合物是葡萄品种中研究最为广泛的一类化合物。在葡萄中发现了大约40种萜烯;它们可以以烃类、醇类、醛类、酮类或酯类的形式存在。这个家族的化合物主要负责果和花香的香气,尽管有一些具有树脂状。一些最具气味的是单萜醇,尤其是芳樟醇(linalool),α-松油醇(α-terpineol),橙花醇(nerol), 香叶醇(geraniol),香茅醇(ci tronel lol)和脱氢芳樟醇( h o t r i e n o l ) 。虽然单萜(monoterpenes)存在于大多数葡萄和葡萄酒中,但在某些品种的马斯喀特和雷司令中尤其普遍。C 13-降异戊二烯(C13-Norisoprenoids)主要来自葡萄中的类胡萝卜素。葡萄中的C 13-降异戊二烯包括β-大马酮(β-damascenone), β-紫罗酮(β-ionone), 3-oxo-α-ionol,β-大马士酮(β-damascone), 和3-hydroxy- β-damascone。萜烯化合物属于植物的次要成分,是由乙酰辅酶A生物合成的,其中间体为异戊烯基二磷酸酯和二甲基烯丙基二磷酸酯。酿酒酵母缺乏具有单萜合酶活性的酶,因此它们不能有效地产生单萜,只有少数天然菌株能够产生少量的单萜。萜烯通过转变成其他化合物而在葡萄酒的品种风味中起着重要作用。
单萜类化合物由两个异戊二烯单元(C10)组成,具有很强的感官潜能。它们是由植物,藻类,丝状真菌和酵母菌从焦磷酸香叶酯生产的。植物组织中存在三种类型的单萜:首先,游离香气化合物如芳樟醇,香叶醇,香茅醇,α-松油醇和肉豆蔻醇等,以及吡喃和呋喃形式的芳樟醇氧化物。还发现了几种单萜醚和乙酸酯。第二,单萜的多羟基化和无味形式,称为多元醇,容易水解具有令人愉悦的味道。第三,单萜的糖苷结合形式,对葡萄汁的香气没有贡献,含量比单萜烯和多元醇的未糖基化形式要丰富。糖苷配基部分通常由萜烯形成,也可能出现其他风味前体,例如直链或环状醇,己醇,苯乙醇,苄醇,C-13去甲肾上腺素,酚酸以及可能的挥发性酚,例如香兰素。
3.8 霉味
食物中最常见的异味之一是霉味或泥土味,通常与微生物的活性有关。导致发霉香气的挥发性化合物包括大约20种化合物,即卤苯甲醚,土臭素,2-甲基异冰片醇,几种甲氧吡嗪,1-辛烯-3-醇,1-辛烯-3-酮,反辛烯醇, 3-辛酮,苯酚和茴香酮。葡萄酒中主要的感官缺陷之一通常与发霉或发霉的香气有关,与软木塞的使用有关。
卤苯甲醚是造成这种霉味的主要化合物。从地窖,软木塞和木桶中分离出来的真菌主要负责由木质素分解产物和氯化化合物生产氯酚。这些最后的化合物源自用于漂白软木塞和洗涤桶的氯化溶液,以及源自橡树林和木材防腐剂中使用的氯化杀菌剂。另外,由于2,4,6-三溴苯酚在地窖中用作杀菌剂,因此可能存在。卤酚可以被细菌转化成它们相应的卤代茴香醚。因此,软木塞中存在2,4,6-三氯茴香醚(2,4,6-trichloranisole,TCA)是造成这种软木塞缺陷的主要原因,其浓度为10–40 ng / L会造成葡萄酒的缺陷(Callejón et al. 2016)。
4 结论
利用遗传操作来增强风味鉴于酵母代谢物对葡萄酒风味的重要性越来越大,研究人员正在探索利用基因手段来生产葡萄酒的可行性也就不足为奇了。有两种方法来处理绒毛的形成,改变囊孢子菌的现有基因和引入新的途径和活动。前一种方法可以包括传统育种或基因工程。引入外源基因需要将这些菌株指定为转基因生物(GMO)。目前,消费者和生产者都对转基因葡萄酒不感兴趣。然而,这种情况在未来可能会改变。现有性状的遗传修饰可能包括在体外产生一个有针对性的等位基因改变,或从另一个菌株或不同种类的酵母菌中分离一个自然发生的特定等位基因,然后在预期的菌株中进行等位基因替换。另外,启动子或终止子序列的改变可以改变基因或蛋白质的表达或调控模式,从而控制有利化合物的形成。构建一个新的杂交蛋白,将显示一个改变的活动或翻译后的调节,也可以用来操纵代谢产物的生产。在某些情况下,可能需要一个空等位基因来消除不需要的活动。然而,无效等位基因可能会对基因组的抑制或其他补偿性改变施加强大的选择压力,并可能导致一个毒株无法与将出现在酿酒厂的酵母菌的野生毒株竞争。上面已经讨论了许多这样的方法。酵母菌菌株表达酶活性,增强葡萄的风味和香气特征也正在开发和测试。