熟期调控岗位
樊绍刚 吴胜 朱明涛 余俊 白描 杨国顺
摘 要:研究葡萄冬芽生理休眠形成和解除分子机理,是研制新的高效无毒休眠解除方法的前提条件。文章简单介绍葡萄芽休眠相关基本概念,然后详细介绍葡萄冬芽生理休眠形成主要原因,光周期、激素、温度和水等因素研究进展情况,探讨生理休眠形成机理。根据提出的休眠解除模型,深入探讨人工破眠处理下生理休眠解除分子响应机理,并归纳研究成果支持论证休眠解除模型。最后探讨葡萄芽休眠研究下一步研究方向。
关键词:葡萄;生理休眠;休眠解除;应激;机理
葡萄(Vitis vinifera L.)芽休眠是适应环境变化的一种生存策略,是茎端分生组织(Shoot apicalmeristem,SAM)为保护自身而随环境变化在活跃生长和休眠状态之间转变的动态过程。芽休眠是一个兼具科学和生产意义的研究课题,对于热带、亚热带暖冬地区和设施栽培葡萄生产尤为重要。长期以来,国内外学者致力于研究芽休眠形成原因、生理特征、品种差异和解除机理,着力解决生产上春季萌芽时间晚、萌芽不整齐,产期调控如促早栽培、延后栽培、二次冬果等问题,从而实现葡萄高效栽培管理,延长鲜果供应期,获得最大经济效益。
葡萄冬芽生理休眠机理研究为生产技术改进提供理论支持,也是开发高效安全休眠解除新方法的首要任务。本篇以Lavee 和 May发表综述文章为基础,总结之后葡萄芽休眠研究进展,重点从生理胁迫级联反应阐述生理休眠解除模型,结合模式植物和其他物种中的相近研究,探索葡萄芽生理休眠形成原因。并对今后研究方向作出展望,以期望开发更为安全高效的休眠调控技术提供理论支持,满足葡萄产业发展多样化需求。
1 芽休眠相关概念
1.1 芽的类型

葡萄芽有两种类型: 夏芽(Prompt bud)和冬芽(Winter bud)。夏芽从最早形成的侧生分生组织发育而来,是真正的腋芽,不经休眠当季萌芽形成副梢。冬芽形成稍晚于夏芽,由自身新梢的顶端分生组织发育而来,位于副梢基部叶腋处。冬芽具有晚熟性,一般要经越冬后次年春季萌发生长,因此称为冬芽或休眠芽(Dormant bud)。冬芽是复芽(Compound bud),也称芽眼, 一般由一个大的主芽(Primary bud)和两至多个小的副芽(Secondary buds)组成,如图1所示。冬芽越冬后部分芽不萌发呈休眠状态,枝蔓基上的小芽随枝蔓增粗潜伏于表皮组织之间,称为潜伏芽或隐芽(Latent bud),冬剪后保留的冬芽是形成下一年产量的主要来源。
1.2 芽休眠定义
葡萄芽休眠是物种为抵御低温、干旱等逆境胁迫进化而来的生理机制。逆境来临时进入休眠维持最低水平的生理状态,环境适宜时恢复生长。芽休眠与种子休眠相比是一种不完全休眠,休眠期生长停止仍有呼吸代谢,但不萌发。
Lang等将芽休眠定义为“包括分生组织在内的任何结构可见生长的暂时停止”。并根据休眠诱导因素将其分为3类:(1)类休眠(Para-dormancy,PD):春夏季节由植物细胞内部特殊的化学信号诱导产生的生长停滞现象,该化学信号不是由休眠组织内部信号独自诱导产生的,而是由植株其他部位器官产生,诱导因素包括顶端优势、日长差异和光周期等。(2)生理休眠(Endodormancy,ED):指植物内部原发性反应所引起的生长停滞现象,这种原发性反应是由休眠组织内部信号独自诱导引发的。(3)生态休眠(Ecodormancy,ECD):冬芽从生理休眠中释放后,由于环境因子限制暂未萌发,当环境条件适宜时即可迅速恢复生长。
这一休眠类型分类方法局限性在于概念中分生组织“可见生长”界定模糊,芽休眠本质上是指生长势停止且不可恢复,植物分生组织生长极其微小或隐藏在芽器官内,不可见并不代表生长停止;分生组织生长包括细胞分裂和细胞伸长,生理活动在时间和空间上独立发生,生长停止难以判断。Lavee等根据葡萄生长周期,参考Lang方法将冬芽休眠分为前休眠(Predormancy)、休眠(Dormancy)和后休眠(Post-dormancy)3个阶段,强调休眠形成和休眠解除之间的动态变化过程。前休眠代表休眠起始,芽在外界环境和诱导刺激下,恢复活跃生长势的可能性逐渐降低。后休眠代表休眠解除的起始,分生组织活跃生长逐步恢复。
2 葡萄冬芽生理休眠形成原因
短日照光周期(Short days,SD)和低温是诱导葡萄进入生理休眠的两个主要环境因子。北半球9月份随温度降低和日照变短,冬芽进入类休眠一段时间后,给予适宜条件其依旧无法迅速萌发即进入生理休眠。首先在枝梢基部节处发生,同一枝梢大概2~3周完成过渡,新梢由绿色变为黄褐色。SAM停止伸长生长且新梢顶端不再形成新的芽眼,茎尖脱落,未完成木质化的副梢开始脱落。伴随进入生理休眠,芽开始形成耐冷性抵御严冬。试验中常用室温条件下萌发响应时间和萌发率BR50区分类休眠和描述生理休眠深度。
2.1 短日照光周期诱导作用
葡萄冬芽进入生理休眠由SD驱动。植物光感应器主要有光敏色素、隐花色素和向光素,通过接收光周期信号感知季节变化,从而调控开花、茎伸长、叶片生长、芽休眠等生理进程。
Sreekantan等发现SD处理延缓但不能停止葡萄花发育进程,可认为葡萄是兼性长日照植物。光敏色素基因VvPHYA和VvPHYB表达受生物钟调节,表达丰度在长日照光周期(Long days,LD)诱导下呈早高晚低规律性变化,而在SD诱导下规律性变化消失,表达量降低。葡萄叶片和冬芽可同时接受光信号,提前落叶延迟生理休眠形成。
开花控制基因FT(FLOWERING LOCUS T)调控细胞周期基因(Cell cycle genes,CCG),SD通过负调控FT表达进入休眠。从PD向ED转变时,叶内和芽内VvFT 同步下调。使用单氰胺(Hydrogen cyanamide,HC)处理后VvFT 上调表达,说明植株负调控VvFT 表达进入休眠,上调VvFT 基因促进芽活跃生长。SD诱导下茎尖组织VvFT 与花分生组织决定基因VvAP1 (APETALA1)、VvAIL2(AINTEGUMENTA-like 2)表达增强, 细胞周期蛋白依赖激酶VvCDKB2 , 细胞周期基因VvCYCA1 、VvCYB 、VvCYD3.2 表达量茎尖较芽内增强,茎尖分生组织通过加速有丝分裂阻止SAM进入生理休眠。SD诱导下叶片和芽内VvFT 均下调表达,HC处理诱导芽内VvFT ,VvAP1 和细胞周期蛋白依赖激酶VvCDKA、VvCDKB2 、细胞周期基因VvCYCA1 上调表达,说明VvFT在调控细胞周期基因转录表达上至关重要,VvFT通过调控细胞周期基因,从而介导生长停止和进入休眠。
SD诱导冬芽进入生理休眠时,芽内出现氧化应激反应,激活细胞抗氧化机制。SD诱导下过氧化氢酶VvCAT、过氧化物酶VvPrx 在低温条件下增强表达水平。Victor等分别用SD和LD诱导萌发后35 d河岸葡萄新梢,发现SD诱导下新梢生长和节位数量显著减少,光呼吸增强,抗坏血酸合成蛋白、PSⅡ和PSⅠ光捕获蛋白复合体、羧化酶等表达丰度差异显著。George等以杂交选育的SD敏感型植株F2-110和SD不敏感型植株F2-040为试验材料,无标签定量鸟枪蛋白质测序发现SD诱导下过氧化物酶、脱氢酶、超氧化物歧化酶大量积累,推断氧化应激相关响应蛋白表达,标志芽进入生理休眠。
光照诱导芽内能量积累。短日光照诱导芽分生组织细胞壁变薄,淀粉合成含量增加,同时低温诱导淀粉降解可溶性糖含量增加,上调脱水素基因表达。SD诱导下ABA可能通过短营养周期蛋白SVP调控生理休眠。Díaz-Riquelme等根据转录组学数据分析,推测开花调控基因VvFLC2 、短营养周期蛋白VvAGL15.1 、VvSPL14-L等基因可能参与生理休眠调控。Fennell等利用SD响应基因型河岸葡萄植株和SD非响应基因型V. hybrid,(Seyval)为材料,分别用SD和LD诱导后分时期取样作转录组和代谢组分析,将基因表达过程分为接收期、诱导期、休眠期3个阶段。ABA、海藻糖、棉子糖等是休眠形成的潜在因素,而在模式植物杨树研究中发现,SD通过激活ABA信号途径诱导休眠。ABA通过SVP同源基因SVL上调胼胝质合成酶CALS1表达,抑制降解胼胝质葡聚糖酶编码基因表达,胞间连丝被胼胝质堵塞,阻隔信号传递,并抑制GA产生,芽组织进入休眠。葡萄休眠冬芽破眠剂处理后葡聚糖酶上调表达,但葡萄SVP基因在冬芽生理休眠的作用还有待验证。猕猴桃中也发现SVP基因在冬芽中表达积累,可能与休眠相关。
2.2 激素在生理休眠中的作用
葡萄内源激素中, 生长素(Auxin,AUX)水平仅与芽生长有关,一般既不参与休眠形成,也不参与休眠解除。细胞分裂素(Cytokinlin,CTK)与花芽分化密切相关。赤霉素(Gibberelli acid,GA)在自然休眠解除过程中具有积极作用,在芽强制起始具有抑制作用,分生组织起始阶段具有抑制作用,保护芽休眠。在起始阶段后可增强原基分化。
脱落酸(Abscisic acid,ABA)与芽生理休眠诱导紧密相关。ABA水平变化主要表现在芽尖,芽鳞中含量极低且基本无变化。内源性ABA含量随休眠深度加深而含量增加,ABA含量达最高点休眠最深。之后ABA含量大幅下降,但休眠深度之后60 d保持不变。休眠是ABA介导的芽分生组织活动的抑制,去除ABA抑制因素即可解除休眠。ABA主要在老龄叶片中合成,秋季人工去叶被认为可有效降低ABA积累,防止休眠发生。ABA在葡萄芽中以游离和糖化形式存在,糖基化与游离ABA比例从休眠期11月份的2倍左右,降低到后休眠期2月份时的37%,而ABA总含量则基本相同。
生理休眠解除时ABA含量降低,糖含量升高,呼吸加剧,细胞分裂加快。芽内糖基化ABA含量随芽含水量增加显著下降,而游离ABA含量基本不变。
外源ABA也可诱导生理休眠。外施适宜浓度ABA可提高耐寒性,叶面喷施高浓度ABA(>800 mg•L-1)产生植物毒害,400~600 mg•L-1抑制枝梢生长,提早暂停茎尖生长和周皮形成,叶片衰老,导致休眠提前。葡萄对外源ABA的反应受叶龄和冷处理影响,ABA对抑制幼龄藤蔓茎生长和增加周皮形成有效,有效诱早枝梢生长停止、叶片衰老和脱落,诱导老藤中100~120 d叶龄老叶进入休眠。
ABA通过抑制细胞周期基因CCG表达进入休眠。Vergara等发现进入ED后ABA合成基因VvNCED1 、VvNCED2 表达水平和ABA含量芽内比茎尖高,而ABA羟化酶基因VvA8H3 则无差异。在生理休眠期间,ABA含量增加,CCG表达下降。ABA可下调芽内和茎尖组织CCG如细胞周期蛋白依赖激酶VvCDKB1 、VvCDKB2 ,A类周期蛋白VvCYCA1、VvCYCA2、VvCYCA3 、B类周期蛋白VvCYCB和D类周期蛋白VvCYCD3.2a ,上调细胞周期蛋白依赖激酶抑制子VvICK5 。而HC处理降低ABA含量,上调CCG表达。同步使用HC和ABA处理冬芽则表现为ABA含量下降,但CCG不上调表达,ABA含量和细胞周期基因活性呈负相关。
ABA介导休眠芽组织内营养积累。ABA含量增加同时淀粉含量也升高。进入休眠后,淀粉依然积累。外施ABA促进单芽插条淀粉含量升高,上调淀粉合酶VvSS1 和VvSS3 ,下调蔗糖代谢基因如转化酶VvINV,蔗糖磷酸合酶VvSUPS等。
2.3 温度和水分作用
低温诱导生理休眠和增加休眠深度。Mathiason等发现低温使参与碳水化合物和能量代谢的基因受抑制,参与信号和细胞生长的基因被激活。低温降低芽体内组织细胞生理活动水平,使芽体进入深度休眠。Perez等比较智利热带和温带两地温度与休眠深度、长度关系,发现在在热带暖冬地区,较高的冬季温度在不改变休眠时间前体下减弱休眠深度,高温增强线粒体呼吸强度改变氧化代谢从而增加H2O2积累水平。在生理休眠末期,热带暖冬地区休眠芽交替氧化酶转录水平较温带下降,线粒体呼吸强度随温度增强等因素使H2O2积累水平较高,休眠冬芽个体间积累差异较大,可能是造成暖冬地区萌芽不整齐的主要原因。
冬芽进入生理休眠期,含水量大幅下降。休眠期冬芽含水量下降到40%且稳定,萌芽前回升至60%附近,萌芽前一周出现伤流现象。浆果发育期间发生水分胁迫可通过延迟生理休眠起始从而缩短休眠周期,减少生理休眠释放所需冷量。干旱胁迫诱导的调控网络在环境与植物激素信号交互中操控生长活动-休眠循环,通过水分胁迫实现活跃生长-休眠周期改变。
3 冬芽生理休眠解除机理
3.1 休眠解除的响应信号
生理休眠状态的芽对环境信号刺激不敏感,条件有利也不萌发,除非出现打破休眠信号。葡萄打破冬芽休眠方法较多,高温、低温、物理或化学等处理方法和刺激均改变休眠芽生理状态。休眠解除和生长恢复信号通路是弄清休眠解除机理关键。
过氧化氢酶(Catalase,CAT)与H2O2是葡萄芽休眠解除的重要信号,对温度和化学刺激反应灵敏,因此成为生理休眠研究的切入点和方向。CAT活性与萌芽率呈负相关,破眠剂诱导可降低CAT活性。CAT活性在10月份达最高值,之后3个月活性随气温下降递减至峰值的1/2。在5 °C条件下HC处理休眠芽3 d后CAT活性降低25%,17 d后降低31%,对过氧化物酶(Peroxidase,Prx)活性则无影响。
Or等鉴定得到第一条葡萄CAT基因序列VvCAT1 ,通过Northern blot验证HC处理后VvCAT1 基因表达丰度迅速降低,说明HC通过抑制CAT基因表达影响CAT活性,而在自然进程中CAT活性无变化。Perez等进一步分离得到3种CAT同工酶,其活性被HC完全抑制。比较分析暖冬和温带地区葡萄CAT活性和H2O2水平之间关系,发现冬季低温(需冷量)和HC作用相同,在于抑制CAT活性。HC处理比自然需冷条件下更早诱导H2O2积累出现峰值和萌芽,因此H2O2可当作自然休眠解除的信号分子。HC处理抑制CAT活性从而使休眠芽内H2O2积累水平上升,外源H2O2同样具有诱导破眠的功能,HC是通过H2O2信号分子介导的休眠解除。
钙信号也参与芽休眠解除。Pang等通过HC诱导对钙传感器表达谱的影响, 发现HC诱导编码钙调素VvCaM ,钙调素结合蛋白VvCBP ,VvCa2+-ATPase 的基因明显上调表达。质膜钙通道阻滞剂LaCl3和钙螯合剂EGTA可抑制HC萌芽诱导,添加外源Ca2+又可消除抑制作用。HC处理增强钙依赖性组蛋白磷酸化程度。
SNF-like蛋白激酶VvGDBRPK可能作为应激反应的信号放大器。HC处理诱导VvGDBRPK上调表达,同时检测到丙酮酸脱羧酶(PDC)和乙醇脱氢酶(ADH) 瞬时诱导,AMP/ATP比值短暂升高,由此推测HC处理迫使植物产生应激反应,AMP作为应激信号传递给SNF-like激酶。进一步分析发现HC诱导上调氧化应激相关基因,如硫氧还蛋白h(TRXh)、谷胱甘肽S-转移酶(GST)、抗坏血酸过氧化物酶(APX)、谷胱甘肽还原酶(GR)。自然低温同样导致类似生理反应,GST、APX、GR和谷胱甘肽过氧化物酶上调表达。
3.2 生理休眠解除模型
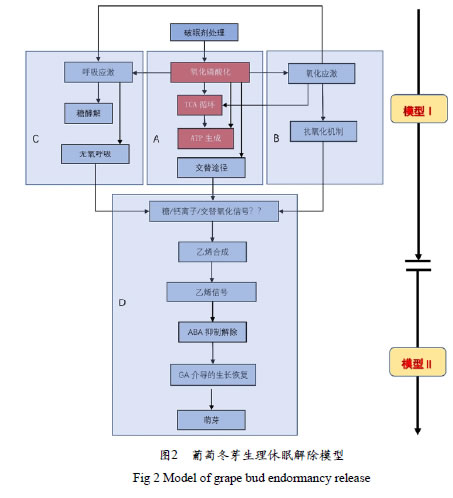
无论是破眠剂诱导还是自然条件下生理休眠解除均是一个极为复杂的包含大量细胞和生化响应过程的调控网络。Keilin等构建HC诱导表达的EST文库共获得5 516条非冗余功能序列,利用比较基因组学发现氧化应激、细胞内囊泡运输、钙信号传导、糖无氧呼吸等生化过程中的功能基因可能参与休眠解生化过程。Halaly等比较HC和热击(Heatt shock,HS)两种处理方式,发现均可解除生理休眠,触发细胞生化水平暂时性呼吸和氧化应激,诱导CAT、ADH和PDC等相关基因瞬时上调表达,破眠处理还诱导抗坏血酸过氧化物酶,谷胱甘肽还原酶、硫氧还蛋白h、谷胱甘肽S-转移酶和蔗糖合成酶编码基因表达。在此基础上,Ophir等利用DNA微阵列技术分析HC和HS诱导表达的EST文库,发现两种处理的应答基因和转录本瞬时调控均存在较大相似性。根据线粒体电子传递链功能受损特征,提出存在激活替代电子转移途径的可能性,并通过原位杂交验证交替氧化酶的存在,其通过稳定线粒体泛醌库氧化还原状态,防止过量活性氧(Reactiveoxygen species,ROS)产生,导致呼吸增加而不伴随ATP产生,进一步提出葡萄休眠芽休眠解除模型(见图2),破眠剂处理下活性氧类产物增加,三羧酸循环活性减弱,ATP产量减少。分子水平表现为氧化应激类基因表达上调,电子转移类基因下调,与糖酵解、丙酮酸代谢、厌氧呼吸途径相关功能基因被诱导上调,大量蛋白质降解。这些都是生物在低氧条件下为应对能源危机选择性诱导的策略,抗氧化机制和相关途径被平行诱导,以应对应激性生理胁迫。氧化和呼吸应激动态变化综合分析表明,芽休眠解除过程中亚致死胁迫、线粒体活性、缺氧条件、乙烯代谢和细胞膨胀之间存在某种特定新联系,可能有助于了解休眠解除机制。
3.3 生理休眠解除模型验证
3.3.1 破眠剂诱导交替途径的激活
HC处理后芽内组织细胞线粒体活动被抑制,细胞色素电子传递链被打断,氧化磷酸化电子传递途径受阻。质子泵无法传递能量,有氧氧化中断不再生成ATP,继而反馈抑制TCA循环,产生能量危机。电子传递途径受阻激活交替途径(抗氰呼吸)旁路,电子传递到交替氧化酶消耗氧分子产生活性氧分子,芽内组织缺氧,产生大量热量,提升冬芽局部温度。人工缺氧环境同样可使线粒体活动受到扰动,产生氧化应激,而抗氰呼吸可促进乙烯(Ethylene,ET)合成。
3.3.2 氧化应激与抗氧化机制激活
线粒体扰动发生不完全还原氧化反应,产生超氧化物负离子、过氧化氢和羟自由基,从而激活抗氧化机制如超氧化物歧化酶、过氧化氢酶、过氧化物酶、谷胱甘肽过氧化物酶等表达。ET和H2O2作为信号分子激活抗氧化酶系统,诱导编码抗氧化酶基因和替代呼吸途径的酶,如交替NADH脱氢酶VvaNDs 、交替氧化酶VvAOXs 基因表达。低氧环境处理休眠冬芽24 h,低氧响应基因VvPDC、VvADH、VvSS,VvnsHB 和VvFT 被强烈诱导。破眠处理诱导ROS相关基因诱导表达,ROS和RNS(NO)物质亚致死水平积累,诱发细胞壁松弛膨大,促进发芽。
HC诱导线粒体外末端氧化酶抗败血酸氧化酶上调表达,抗败血酸-谷胱甘肽循环系统高效运转,还原性/氧化型比例上升(GSH/GSSG)。破眠产生的缺氧和呼吸应激,激活γ-氨基丁酸GABA旁路基因表达,造成琥珀酸盐、Ala,GABA积累。破眠处理产生的缺氧和呼吸应激上调GABA旁路基因VvGAD1 表达,但GABA量保持不变,通过琥珀酸盐纳入TCA循环,避免GABA积累毒害。外源GABA抑制抗氧化酶APX、GSH-Px、SOD、CAT1表达,但上调CAT2表达,从而降低破眠处理诱导的H2O2水平。
3.3.3 呼吸应激下的物质和能量代谢
呼吸和氧化应激条件下可溶性糖迅速耗竭,诱导胞浆内糖酵解反应快速供能,液泡内磷酸戊糖途径PPP活动增强,产生核糖和NADPH,NADPH是多种合成代谢的供氢体,维持谷胱甘肽还原状态。低氧环境下6-磷酸葡萄脱氢酶VvG6PD,1,3-葡聚糖酶VvGLU上调表达。需冷量不足导致萌芽率低,可能是由于己糖供应量低。糖酵解和乙醇发酵增加可能导致休眠解除机制中下游阶段的激活。
HC处理产生的缺氧导致冬芽内淀粉水解和α-淀粉酶Vv α-AMY1、Vvα-AMY2、Vvα-AMY4表达上调,可溶性糖在芽和节间组织中短暂积累。可溶性糖(葡萄糖和果糖)作为低氧条件的传递信号,通过低氧效应改变AMY基因表达。赤霉素诱导AMY3和AMY4表达,破眠处理则诱导AMY1和AMY2表达, 两种酶与芽的生理休眠解除相关。HC处理后芽组织内蔗糖、葡萄糖和果糖浓度迅速下降,而酸转化酶活性在处理过的芽中保持较高,但在节间却没有。芽可利用强大的输导组织从底层组织中吸引蔗糖,从而导致细胞代谢重新激活,蔗糖可能起信号的作用。
3.3.4 破眠信号传导
破眠信号传导尚无明确途径,根据当前研究结果,认为破眠剂诱导生理休眠解除过程可能通过一种不具有特征的生化级联而导致休眠解除。
3.3.5 ET和ABA代谢互作
生理休眠是通过ABA介导的抑制芽分生组织活动维持生长停止状态,消除ABA抑制作用即可触发休眠释放,外源ABA还可使HC破眠效果衰减。HC处理下调ABA合成关键酶基因VvXERICO 和VvNCED1转录水平,并诱导ABA降解酶ABA8'-羟化酶VvA8H-CYP707A4 大幅上调表达,ABA含量降低。ABA 8'-羟化酶自然条件在从生理休眠向休眠解除的过程中实现量的积累,超表达VvA8H-CYP707A4 发现转基因植株表现出较高的ABA分解代谢能力,显著提高萌芽率,并得到更长的夏季基部侧梢。乙烯诱导冬芽生理休眠释放,Shi等证实,破眠剂处理诱导乙烯生物合成,外源乙烯具有解除休眠的作用,抑制乙烯生物合成可衰减破眠剂作用。休眠解除前乙烯合成短暂增加主要通过调节特定ACC合成酶(ACC synthase,ACS)和ACC氧化酶(ACC oxidase,ACO)基因转录实现,破眠处理信号刺激显著诱导VvACS1、VvACS6、VvACO2 上调,ET水平随之发生变化。在生理休眠解除过程中,ACC氧化酶转录产物和乙烯水平急剧下降,而ACC在自然休眠过程中积累。
3.3.6 分生组织生长恢复
GA介导休眠解除生长恢复,GA4对生理休眠解除有促进作用。Zheng等发现,在生理休眠诱导过程中,GA活性转化基因VvGA3ox(转化GA12为GA4))、VvGA20ox(转化GA12为GA1)和具有生物活性的GA1含量水平下降。在休眠解除过程中,基因表达增强,且伴随GA失活基因VvGA2ox 表达减弱,GA1含量升高。尽管GA在自然休眠释放过程中起积极作用,但GA在芽破眠剂诱导时同步使用时具有抑制作用,但在分生组织激活后,GA水平增加有助于促进原基重新生长。破眠处理增加生长素合成关键基因VvYUC3 、CTK生物合成相关基因VvIPTs 和VvLOG1 表达及降低CK分解代谢相关基因VvCKX3 表达,增加CK水平,萌芽之前CK和生长素水平增加。
4 展望
总的来看,近20多年来在该领域的研究上已取得较大进步,特别是2007年葡萄基因组测序结果公布后,组学技术发展,研究人员利用转录组学、蛋白组学、代谢组学等从宏观尺度上探索和揭示冬芽休眠这一复杂生理活动分子机理。当然,这一领域还存在诸多未解之谜,还有很多相似基因功能有待验证,我们认为,今后葡萄芽休眠研究应集中在以下方面:
(1)冬芽生理休眠形成机理的深入
在模式杨树植物中发现,SD激活ABA合成通路,ABA通过上调胼胝质合成酶CALS,抑制葡聚糖酶活性,阻断胞外信号传导路径,下调GA信号通路基因表达,进入休眠。在葡萄中,ABA与SVP及下游胼胝质合成酶CALS,GA调控机制还有待研究。
(2)休眠解除模型进一步验证
参照休眠解除模型,需冷量可看作是低温环境下冬芽组织缓慢积累ROS以达亚致死胁迫的时间长度,HC处理和满足需冷量自然条件下,从呼吸应激和氧化应激反应到ET与ABA代谢互作、GA介导的生长恢复信号通路和调控机制知之甚微,例如梨miR6390参与降解DAM基因而解除休眠。有关葡萄休眠积累的miRNA研究还有待进一步深入。