南宁综合试验站
成果 张劲 黄小云 张瑛 谢太理 谢林君 余欢 周咏梅 周思泓
摘 要:【目的】初步分析广西特色酿酒葡萄品种‘桂葡6号’和‘凌丰’葡萄黄烷-3-醇组分特征,为今后系统开展特色酿酒品种酚类物质代谢相关研究奠定方法基础和提供理论参考。【方法】本研究以不同发育期毛欧杂种‘凌丰’和野生资源选育的‘桂葡6号’葡萄为试验材料,利用高效液相色谱(HPLC)技术检测其果皮和种子中黄烷-3-醇类物质的含量,并与欧亚种‘赤霞珠’比较。【结果】浆果整个发育期中,‘桂葡6号’葡萄果皮中黄烷-3-醇总量显著高于‘赤霞珠’和‘凌丰’;浆果成熟期,‘桂葡6号’和‘赤霞珠’种子中黄烷-3-醇显著高于‘凌丰’;葡萄果皮和种子中黄烷-3-醇总量、儿茶素、表儿茶素、表棓儿茶素及表儿茶素没食子酸酯含量随发育期的变化趋势在不同品种间存在差异;果实成熟后期,‘凌丰’和‘桂葡6号’葡萄果皮中3’5’-羟基取代黄烷-3-醇比例高于‘赤霞珠’,而整个发育期‘桂葡6号’种子中3’5’-羟基取代黄烷-3-醇比例显著高于其它两个品种。【结论】野生资源选育的‘桂葡6号’果皮和种子中黄烷-3-醇含量和3’5’-羟基取代黄烷-3-醇的比例较高,说明‘桂葡6号’黄烷-3-醇酚类化合物含量丰富,在提高葡萄酒柔顺度和护色方面具有良好品质。
关键词:‘桂葡6号’;‘凌丰’;果皮;种子;黄烷-3-醇
【研究意义】目前,我国酿酒葡萄生产中存在应用品种较为单一的问题,仍然以欧亚种葡萄‘赤霞珠’、‘霞多丽’等为主。而广西属亚热带气候区,高温多湿,欧亚种葡萄品种由于抗病性较差一直未在当地广泛种植。南方野生葡萄品种耐高温高湿,抗病性强,不易感染黑痘病、白腐病、炭疽病等病害,是重要的葡萄品种资源。因此,挖掘广西本地葡萄品种资源,提升特色酿酒葡萄品质至关重要。【前人研究进展】原花青素(Proanthocyanidins,Pas), 又称为缩合单宁(Condnsed Tannins,CTs),在植物体经苯丙烷-类黄酮代谢路径合成,来源于葡萄果皮和种子,构成葡萄酒的‘骨架’,并保证酒的色泽稳定性。原花青素是黄烷-3-醇(Flavan-3-ol)单元的聚合体,葡萄中常见的黄烷-3-醇单体有:儿茶素(Catechin, C)、表儿茶素(Epicatechin,EC)、表棓儿茶素(Epigallocatechin,EGC) 、表儿茶素没食子酸酯(Epicatechingallate,ECG)。葡萄果实成熟过程中黄烷-3-醇组分含广西两个特色酿酒葡萄品种黄烷-3-醇组分解析量发生变化。研究表明,不同葡萄品种单宁含量一般在坐果时达到最高,进入转色期时含量开始下降。
不同品种、栽培条件、紫外线照射、激素处理等对葡萄果皮和种子中黄烷-3-醇物质的组成和含量有很大影响。前人研究表明,北美种群和欧美杂种葡萄中黄烷-3-醇含量最高,圆叶葡萄次之,而多数东亚种葡萄中黄烷-3-醇含量较低。UV-C 能够增强隐色花色素还原酶活,从而诱导葡萄果实中黄烷-3-醇的合成。李舒婷等人(2018)研究表明,避雨栽培虽然对黄烷-3-醇总量没有显著影响,但提高了甲基化黄烷-3-醇比例。高翔等人(2014)研究表明,油菜素内脂处理能够提高‘赤霞珠’果皮及种子中的黄烷-3-醇含量。【本研究切入点】近年来国内外学者不断改进葡萄果实及葡萄酒中黄烷-3-醇类物质的检测方法,并探究不同品种、不同产区葡萄及葡萄酒黄烷-3-醇类的组成及含量差异,而此类研究多集中于欧亚种葡萄。针对广西特色酿酒葡萄品种‘凌丰’和‘桂葡6号’中黄烷-3-醇类物质组分特征的研究未见报道。【拟解决的关键问题】以广西特色酿酒葡萄品种‘凌丰’和‘桂葡6号’为试材,并与欧亚种‘赤霞珠’进行比较,通过检测冬果不同发育期果皮和种子中黄烷-3-醇的含量,分析其组分特征,为今后优化广西酿酒葡萄的栽培措施,提高当地特色酿酒葡萄的品质提供理论基础。
1 材料与方法
1.1 试验地点与材料
试验地位于本研究所基地葡萄园(22°50′59″ N,1 0 8 ° 1 4 ′ 3 5 ″ E ) 。供试的葡萄(Vi t i s L . ) 品种为‘ 赤霞珠’(V. vinifera L.)、‘桂葡6号’(Vi t i s s p .)以及‘凌丰’(V.quinquangularis ×V.vinifera ),树龄分别为8年、3年和10年。三个品种的整形方式分别为:单干双臂型、一字型和Y字型,行距2.5m,株距1.5 m,南北行向,灌溉方式均为滴灌。三个品种果实样品均为冬果,采集时间始于花后4周(2015年9月28日),之后每两周采样一次至采收期结束。每个品种选择树体长势基本一致的9株作为实验树,每3株合并作为一个生物学重复。每次采样时间为上午9:00~10:00,样品采后放入冰盒立即运回实验室。
1.2 果实理化指标测定
从每个生物学重复的实验树上剪下100粒浆果果粒(即每个采样点每个品种共300粒),随机兼顾果穗的各个部位。其中150粒样品(每个生物学重复50粒)用于测定果实鲜重、单粒果皮鲜重、单粒种子数量、单粒种子鲜重、可溶性固形物含量和可滴定酸含量。详细步骤如下:果实称重后破碎挤汁,然后将果皮、果肉、种子分离,滤纸吸去果汁后称量果皮和种子鲜重,并统计种子的数量。利用PAL-1型手持折射(Atago,Tokyo,Japan)测量可溶性固形物(Total solubles o l i d s ,TSS) 含量。可滴定酸(Titratable Acid, TA)含量利用酸碱滴定法进行测定,结果用酒石酸表示。每个生物学重复采取3个技术重复测定。剩余样品用液氮速冻后保存于-80℃冰箱待用。
1.3 黄烷-3-醇类物质的检测
黄烷- 3 - 醇的提取方法参照Liang等的方法。取0.1g葡萄皮干粉或种子干粉于10mL离心管中,加入1mL间苯三酚缓冲液(含0.3NHCl和 50g/L 间苯三酚)和0.005g抗坏血酸,在50℃下静置裂解 20min后,加入1mL乙酸钠(200mM)终止反应,离心15min (10000rpm)后取出上清液置于新的 10mL离心管中;重复提取3次合并上清液摇匀,于-80℃保存待用。每个生物学重复单独提取。
采用岛津LC-20AD高效液相色谱对提取产物进行检测。流动相A: 0.2%乙酸水溶液,流动相B:乙腈:0.2%乙酸水溶液=4: 1。色谱条件如下:梯度洗脱程序:0~20min,10.0%B;20~30min,10~15%B;30~40min,15~20%B;40~50min,20~33%B;50~55min,33~40%B55~58min40~100%B;58~63min,100%B;63~64min,10%B。流速:1.0mL/min;柱温:25℃;检测波长:280nm;进样量:25μL。
1.4 葡萄果皮和种子中黄烷-3-醇类物质的定性与定量
本研究的黄烷-3-醇定性工作对标准品的保留时间一一确定。分别建立1~1000mg/L之间、9个水平、三个重复的儿茶素、表儿茶素、表棓儿茶素和表儿茶素没食子酸酯标准曲线,相关系数在0.97以上。果皮和种子中黄烷-3-醇含量单位用μg•浆果-1表示。
1.5 试剂与标样
标准品: 儿茶素[ ( + ) - C a t e c h i n ] 、表儿茶素[(-)-Epicatechin]、表棓儿茶[(-)-Epigallocatechin]和表儿茶素没食子酸酯[(-)-Epicatechingallate]均购买于 Sigma(St. Louis, MO,USA)公司,纯度均98%以上。
色谱纯试剂: 乙腈( A c e t o n i t r i l e ) 、甲醇(Methanol)和乙酸(Acetic acid)均购买于Fisher(Fairlawn, NJ,USA)公司,纯度均达99.9%以上。
分析纯试剂: 间苯三酚(Phloroglucinol)购买于上海瑞永生物科技有限公司;抗坏血(Ascorbate)购买于广州金华大化学试剂有限公司;乙酸钠购买于天津博迪化工股份有限公司;浓盐酸(HCl)购买于廉江市爱廉化学试剂有限公司。
1.6 统计分析
利用独立样本T检验在P<0.05水平下进行显著性分析。黄烷-3-醇含量用μg•浆果-1表示。数据处理采用SPSS 20.0(SPSS Inc., Chicago,IL, USA)。绘图采用MicrosoftExcel 2010。
2 结果与分析
2.1 不同品种果实理化指标随发育期变化情况
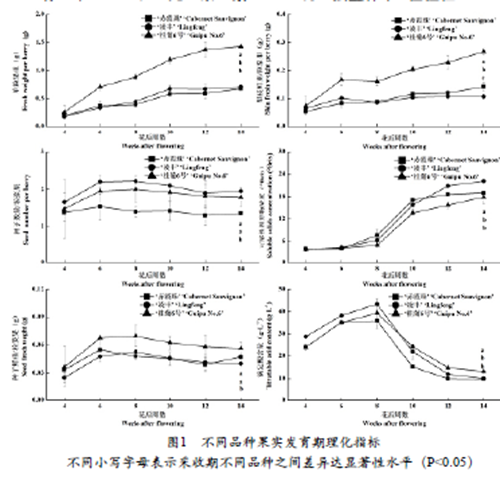
由图1可以看出,三个供试葡萄品种理化指标随发育期的变化趋势基本一致,但是含量高低、数量大小存在差异。‘赤霞珠’和‘凌丰’葡萄果实于花后8周开始转色,而‘桂葡6号’于花后10周进入转色期。自花后6周起,‘桂葡6号’果粒鲜重和果皮鲜重高于‘赤霞珠’和‘凌丰’。采收期(花后14周),‘桂葡6号’果粒鲜重和果皮鲜重约为其它两个品种的2倍。花后6至14周,‘赤霞珠’平均每粒浆果中的种子数量低于‘凌丰’和‘桂葡6号’,且‘桂葡6号’平均每粒浆果中的种子重量高于‘赤霞珠’和‘凌丰’。花后8至14周,‘桂葡6号’果实可溶性固形物浓度始终低于‘赤霞珠’和‘凌丰’。花后12至14周,‘凌丰’果实可溶性固形物浓度最高。花后4至8周,‘凌丰’果实可滴定酸含量最高,但在采收期(花后14周)‘桂葡6号’果实可滴定酸含量显著高于其它两个品种。
2.2 不同品种葡萄果皮黄烷-3-醇含量变化
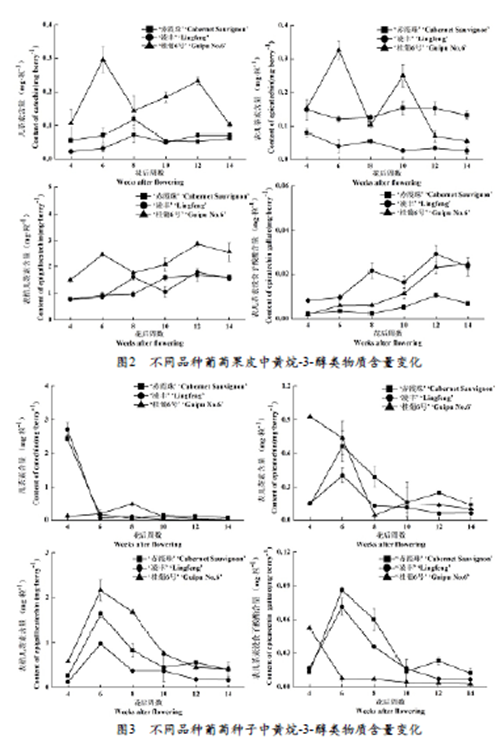
如图2所示,不同品种葡萄果皮中黄烷-3-醇含量随发育期的变化趋势不同。总体来说,三个品种葡萄果皮中表棓儿茶素的含量最高,其次是儿茶素和表儿茶素,表儿茶素没食子酸酯的含量最低。自花后4周至采收(花后14周),‘桂葡6号’葡萄果皮中儿茶素的含量高于其它两个品种,且在花后6周时其含量达峰值,而‘赤霞珠’和‘凌丰’则是在花后8周时含量最高。三个品种葡萄果皮中表儿茶素含量随发育期变化呈波动下降趋势。花后4周至采收(花后14周),‘凌丰’葡萄果皮中表儿茶素含量始终低于其它两个品种。花后12至14周,‘赤霞珠’葡萄果皮中表儿茶素含量最高。花后4至12周,三个葡萄品种葡萄果皮中表棓儿茶素含量呈波动上升趋势,但在采收期(花后14周)又略有下降。‘桂葡6号’葡萄果皮中表棓儿茶素含量始终高于其它另两个品种。在整个发育阶段,‘凌丰’葡萄果实中表儿茶素没食子酸酯含量最高,其次是‘桂葡6号’,‘赤霞珠’含量始终最低。‘桂葡6号’葡萄果皮中表儿茶素没食子酸酯含量在整个发育阶段呈上升趋势,而‘赤霞珠’和‘凌丰’其含量自花后4至12周不断增加,到采收期略有降低。
2.3 不同品种葡萄种子黄烷-3-醇含量变化
如图3所示,不同品种葡萄种子中黄烷-3-醇含量随发育期的变化趋势也存在差异。总体来说,三个品种葡萄种子中表棓儿茶素的含量最高,其次是表儿茶素,再次是儿茶素,表儿茶素没食子酸酯的含量最低。自花后4周至采收(花后14周),三个品种葡萄种子中儿茶素含量在波动中下降,采收期含量将至最低,‘赤霞珠’种子中儿茶素含量自花后10周起高于其它两个品种。花后4周,‘桂葡6号’种子中表儿茶素含量最高,而‘赤霞珠’和‘凌丰’种子中表儿茶素含量在花后6周达峰值后在波动中下降。采收期(花后14周)‘赤霞珠’种子中表儿茶素含量高于‘桂葡6号’和‘凌丰’。三个品种葡萄种子中表棓儿茶素含量随发育期的变化趋势一致,且都在花后6周时含量最高,随后持续降低。花后4至10周,‘桂葡6号’种子中表棓儿茶素含量最高,采收期时‘凌丰’种子中表棓儿茶素含量低于其它两个品种。花后4周时,‘桂葡6号’种子中表儿茶素没食子酸酯含量最高,而‘赤霞珠’和‘凌丰’种子中其含量在花后6周达峰值。花后6至14周,‘桂葡6号’种子中表儿茶素没食子酸酯含量最低,采收期‘赤霞珠’中其含量高于其它两个品种。
2.4 不同品种葡萄果皮及种子黄烷-3-醇总量及比例变化
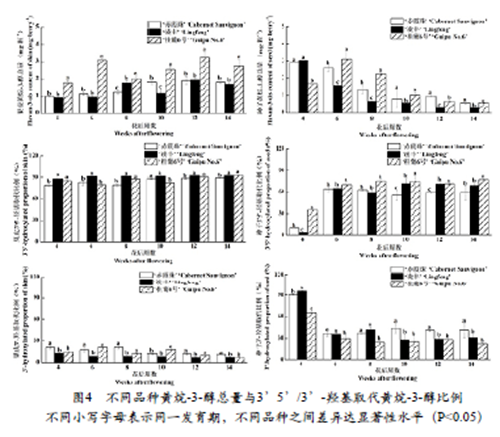
由图4可以看出,三个品种葡萄果皮中黄烷-3-醇总量在花后12周达峰值,且整个发育阶段‘桂葡6号’果皮中黄烷-3-醇总量显著高于‘赤霞珠’和‘凌丰’,浆果成熟后期(花后12~14周)‘赤霞珠’和‘凌丰’之间差异不显著。三个品种葡萄种子中黄烷-3-醇总量随发育期的变化趋势与果皮不同。‘赤霞珠’和‘凌丰’种子中黄烷-3-醇在花后4周含量最高,而‘桂葡6号’则是在花后6周达峰值。进入转色期后,三个品种葡萄种子中黄烷-3-醇总量持续降低,且‘凌丰’种子中黄烷-3-醇总量显著低于‘桂葡6号’和‘赤霞珠’。将黄烷-3-醇按其B环羟基化程度不同分类,可分为3’5’-羟基化和3’ -羟基化两种。本研究结果表明,三个品种葡萄果皮中所含有的黄烷-3-醇以3’5’-羟基化修饰为主,且3’5’-羟基化黄烷-3-醇比例随发育期呈波动上升趋势,而3’-羟基化黄烷-3-醇比例持续降低。花后12~14周,‘赤霞珠’果皮中3’5’-羟基化黄烷-3-醇比例显著低于‘凌丰’和‘桂葡6号’。花后4周,三个品种葡萄种子中所含的黄烷-3-醇以3’-羟基化修饰为主,随后3’5’-羟基化修饰比例急剧增加,到采收期为止,其所占比例>50%。花后4~14周,‘桂葡6号’葡萄种子中3’5’-羟基化黄烷-3-醇比例显著高于‘赤霞珠’和‘凌丰’。果实成熟阶段(花后10~14周),‘赤霞珠’葡萄种子中3’-羟基化黄烷-3-醇比例显著高于其它两个品种。
3 讨论
葡萄果实中常见的黄烷-3-醇类物质有儿茶素、表儿茶素、表棓儿茶素和表儿茶素没食子酸酯四类,它们存在于果实的果皮、种子、果梗、果肉及枝条中。本研究对‘赤霞珠’、‘凌丰’和‘桂葡6号’不同发育期理化指标的检测结果表明,野生资源选育的品种‘桂葡6号’单粒浆果的果皮重量和种子重量高于其它两个品种,而葡萄皮和种子所含的黄烷-3-醇类物质为葡萄及葡萄酒的酚类物质的主要来源,因此野生资源选育的‘桂葡6号’酚类物质含量丰富。
不同品种葡萄中黄烷-3-醇物质的含量有很大差异。Jin et al 对9个红色酿酒葡萄果皮中的酚类物质的检测结果表明,‘西拉’、‘蛇龙珠’‘赤霞珠’、‘美乐’和‘烟73’果皮中的黄烷-3-醇总量显著高于‘佳美’和‘黑比诺’。Zhu etal 对6个种群的23个葡萄品种果皮中的黄烷-3-醇类物质的检测结果发现,北美种群的‘尼亚加拉’果皮中黄烷-3-醇总量最高,约为欧亚种‘赤霞珠’的20倍左右。在本研究中,野生资源选育的‘桂葡6号’葡萄果皮中黄烷-3-醇总量显著高于欧亚种‘赤霞珠’和毛欧杂种‘凌丰’。张娟等对20个红色酿酒品种葡萄种子中黄烷-3-醇类物质的检测结果表明,‘沃波尔特’、‘法国兰’和‘粉红沙斯拉’3个品种籽中黄烷-3-醇总量最高,‘美乐’、‘灰比诺’、‘赤霞珠’和‘佳丽酿’等品种次之;‘黑虎香’、‘北醇’、‘神索’和‘蛇龙珠’最低。在本研究中,果实成熟阶段‘凌丰’种子中黄烷-3-醇总量显著低于‘赤霞珠’和‘桂葡6号’。
黄烷醇类物质除了决定葡萄酒的苦味及涩味以外,还对葡萄酒颜色具有保护作用,由葡萄果皮及种子中经发酵过程进入葡萄酒中。因此,广西本地野生资源选育的‘桂葡6号’在黄烷-3-醇对葡萄酒的护色方面具有很大潜力。
已有大量文献对葡萄生长发育过程中黄烷-3-醇类物质的变化进行了报道,它们在果实的不同发育阶段和不同部位进行合成。葡萄果皮中的黄烷-3-醇在转色后1~2周达到最大值,随后呈急剧降低趋势,主要原因是转色后类黄酮代谢路径上的底物更多的流向合成花色苷的方向。在本研究中,‘赤霞珠’、‘凌丰’和‘桂葡6号’果皮中黄烷-3-醇含量都在花后12周出现峰值,这主要取决于果皮中含量最高的表棓儿茶素的变化趋势。在葡萄种子中,黄烷-3-醇在果实转色前完成积累,转色后到成熟此类物质的含量有明显下降,主要原因是氧化作用。在本研究中,三个品种葡萄种子中黄烷-3-醇总量都在转色前达到峰值,‘赤霞珠’和‘凌丰’种子中黄烷-3-醇总量在花后4周最高,而‘桂葡6号’中其最大值则出现在花后6周。由此可见,不同葡萄品种果皮和种子中黄烷-3-醇随发育期的积累和变化规律存在差异,这对于其生物学性状的描述和酿酒品质的评价具有重要意义。
表儿茶素与儿茶素虽然只是在C3位的构象不同,但却有截然不同的口感,表儿茶素比儿茶素更加苦涩而且苦涩的感觉持续的时间也更长。果实成熟阶段,‘赤霞珠’果皮和种子中表儿茶素含量高于‘桂葡6号’和‘凌丰’,说明其苦涩感比其它两个品种更强。在植物体内,原花色素由黄烷-3-醇单体聚合而成。原花色素结构多变,这主要是黄烷醇的修饰类型、化学异构以及聚合度的变化造成的。黄烷-3-醇B环羟基化模式取决于细胞色素P450单加氧酶F3’H和F3’5’H的催化,由F3’H催化产生的二羟基化(3’-羟基取代)黄烷-3-醇主要有儿茶素和表儿茶素;由F3’5’H催化产生的三羟基化(3’5’-羟基取代)黄烷-3-醇主要有 儿茶素和表棓儿茶素。
果实成熟后期,‘凌丰’和‘桂葡6号’葡萄果皮中3’5’-羟基取代黄烷-3-醇比例高于‘赤霞珠’,而整个发育期‘桂葡6号’种子中3’5’-羟基取代黄烷-3-醇比例显著高于其它两个品种。另外,黄烷-3-醇C3位的羟基基团容易被没食子酸酯化,这类酯化的黄烷-3-醇大量存在于葡萄籽中。没食子酸酯化使酒的单宁感变粗糙,而三羟基化黄烷醇能够降低这种粗糙的口感,柔顺酒体。由以上分析可以看出,‘桂葡6号’果实中高比例的3’5’-羟基取代黄烷-3-醇有助于提高葡萄酒的柔顺度。
4 结论
葡萄果皮和种子中黄烷-3-醇总量、儿茶素、表儿茶素、表棓儿茶素及表儿茶素没食子酸酯含量随发育期的变化趋势在不同品种间存在差异影响。广西特色酿酒葡萄品种‘桂葡6号’和‘凌丰’葡萄果皮和种子中黄烷-3-醇含量最高的一类是表棓儿茶素。浆果成熟后期‘凌丰’种子中黄烷-3-醇总量显著低于‘桂葡6号’和‘赤霞珠’。野生资源选育的‘桂葡6号’果皮和种子中黄烷-3-醇含量和3’5’-羟基取代黄烷-3-醇的比例较高,是极具发展潜力的红色酿酒品种。