生态与土壤管理岗位
耿庆伟 翟 衡 杜远鹏
摘 要:以盆栽‘赤霞珠’葡萄为试验材料,利用叶绿素荧光动力学技术,研究了(120±20)nL•L-1的臭氧浓度下不同光强(800,1600 μmol•m-2•s-1)与温度(26,40 ℃)处理对光系统II性能的影响。结果发现:臭氧胁迫下40 ℃的高温、1600μmol•m-2•s-1的强光均可以显著降低叶片最大光化学效率(Fv/Fm)、线性电子传递速率(ETR)、单位面积有活性反应中心的数量(RC/CSm)以及光化学淬灭系数(qP),同时降低了光系统 I 激发能分配系数(α),增大了光系统 II 激发能分配系数(β),导致两个光系统之间的激发能分配严重偏离平衡,造成了光系统 II 的伤害,但1600 μmol•m-2•s-1的强光影响更为显著;本研究结果表明:田间温光逆境增加了PSII活性对臭氧胁迫的敏感性,其中以强光胁迫的作用效果更显著,而高温强光胁迫下臭氧对PSII活性抑制程度最大。
关键词:葡萄,臭氧,光照,温度,叶绿素荧光参数
近几十年来O3污染已经成为对流层中首要污染物,对作物生长和农业生产造成很大影响,全球近1/4的国家或地区面临O3浓度超出60nL•L-1的威胁。由于人类活动和生物源排放,O3浓度仍以每年约0.5% ~ 2.5%的速率增长,预计2015~ 2050年全球地表O3浓度将在现有基础上增加20% ~ 25%,21世纪末将增加40% ~ 60%。O3具有强氧化性,可对植物产生氧化伤害。
光是植物进行光合作用所必须的环境因子,但在强光下,当光合机构吸收的光能超过光合作用的利用量时,即出现光抑制,若植物不能及时有效地利用或耗散,就会产生过量的活性氧,破坏光合色素和 Dl 蛋白等,引起 PSⅡ的损伤。高温胁迫会破坏叶片PSⅡ,导致植物叶片不能有效利用吸收的光能,最终会降低植物的光合作用、损坏细胞膜并打破活性氧平衡。自然条件下,夏季强光和高温往往同时出现,伴随着臭氧的污染,植物遭受臭氧、强光、高温等三种逆境因子的交叉胁迫。大量气室试验表明,O3胁迫对植物光合作用和产量有所影响,但这种影响是否随着高温、强光等逆境因子的出现而加剧尚不清楚。
本试验中以广泛栽培的酿酒葡萄品种‘赤霞珠’为试材,利用叶绿素荧光技术,探究光合机构的运转状况,探测和分析植物的光合功能,了解臭氧胁迫下强光、高温胁迫对葡萄叶片PSⅡ活性的影响,以期探寻减轻光抑制和提高逆境光合能力的有效途径。
1 材料与方法
1.1 试验材料和处理
试验于2016年7月 ~ 9月在山东农业大学葡萄园内进行。试材为长势一致的‘赤霞珠’( Vitis viniferaL.‘Cabernet Sauvigon’)一年生苗,种植于直径25cm、高35cm的花盆中。基质:沙 = 1:1。待植株长至10 ~ 12片完全展开叶时进行试验处理。
处理分为 5 组,分别在自然大气臭氧浓度条件下正常光照与适温(35 ppb,800 μmol•m-2•s-1,26℃) 、臭氧胁迫下正常光照与适温(120±20ppb,800μmol•m-2• s -1,26℃)、臭氧胁迫下正常光照与高温( 1 2 0 ± 2 0 p p b ,800μmol•m-2 •s-1,40 ℃)、臭氧胁迫下强光与适温(120±20 ppb,1600μmol•m-2•s-1,26 ℃)和臭氧胁迫下强光与高温(120±20 ppb,1600μmol•m-2•s-1,40 ℃)条件下处理 3h(5 个处理分别表示为对照Control、T1、T2、T3、T4),处理结束后进行相关参数测定,每个处理重复 5 次。
1.2 测定项目和方法
采用英国Hansatech公司的FMS-2型便携脉冲调制式荧光仪测定荧光参数,测定程序如下:首先对叶片进行30min暗适应,用饱和脉冲光(12000μmol•m-2•s-1),测定暗适应3 0mi n 后最大荧光(Fm)。对光适应下的叶片先用60s作用光(1000μmol•m-2•s-1),然后用极弱的(<0.05μmol•m-2•s-1)测量光测得叶片最小荧光(Fo'),再用饱和脉冲光(12000μmol•m-2•s-1),测定光适应下的最大荧光值Fm',打开作用光测定光下稳态荧光Ft。包括:天线转化效率:(Fv'/Fm')=(Fm'–Fo')/Fm';线性电子传递速率:(ETR)=(ФPSII)×PFD×0.84×0.5,其中PFD是被吸收的光通量密度(μmol•m-2 •s-1),0.5代表光能在两个光系统间的分配系数,0.84指入射到叶片表面的光能平均有84%被叶片吸收(Genty et al.,1989);光化学淬灭:(qP)=(Fm'-Fs)/(Fm'-Fo')。两个光系统之间的激发能分配系数按Braun等[Braun]的公式计算:光系统Ⅰ激发能分配系数α =f /(1 + f), 光系统Ⅱ激发能分配系数β= 1/(1 + f), f =(Fm'-Fs )/(Fm'-Fo');光系统Ⅰ和Ⅱ间激发能分配的不平衡性可用β/α-1表示。根据 Demmig-Adams等[Demmig-Adams]提出的公式计算光合机构吸收的光能用于光化学反应的份额P =Fv'/Fm' × qP,用于天线热耗散的份额(D)= 1 - Fv'/Fm',用于反应中心由非光化学反应耗散的份额(过剩光能,X 耗散)(Ex)= Fv'/Fm'×(1 - qP)。
用连续激发式荧光仪(HandyPEA,Hansatech,英国)测量荧光诱导动力学曲线,参考李鹏民等(李鹏民 等,2005)的计算方法,获得如下参数:PSII最大光化学效率Fv/Fm,PSII的潜在活性Fv/Fo,单位面积有活性反应中心数目(RC/CSm)=Fm•φPo•(Vj/Mo),捕获的激子将电子传递到QA以后的其它电子受体的概率(Ψo)=ETo/TRo=(1–Vj)。
1.3 数据处理及统计分析
采用Microsoft Excel软件处理数据和制图,采用DPS软件的LSD法进行差异显著性检验。
2 结果与分析
2.1 臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片Fv/Fm、Fv/Fo、RC/CSm、Ψo的影响
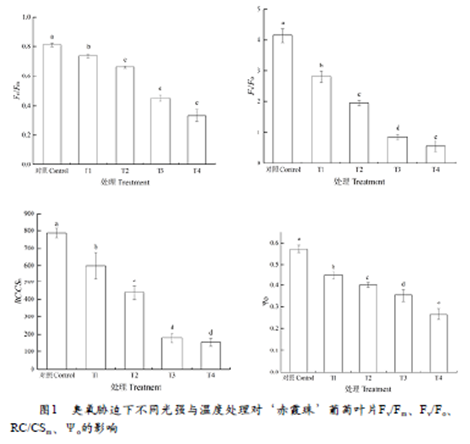
Fv/Fm 是 PSⅡ最大光化学量子产量,或称最大 PSⅡ的光能转换效率,代表了光合机构把吸收的光能用于光化学反应的最大效率,反映光能吸收转化机构的完整性,非胁迫条件下该参数的变化极小,不受物种和生长条件的影响,常用Fv/Fm下降程度来反映光抑制程度的大小。从图1可以看出,与对照相比,不同处理显著降低叶片的Fv/Fm值,与T1相比,T2、T3分别降低了10.3%和38.8% ,T4降幅最大,达到54.8%。Fv/Fo表示的是PSII的潜在活性,与T1相比,T2、T3和T4分别降低了和30.4%、69.6%和80.3%。RC/CSm反映了叶片单位面积内有活性反应中心的数量,Ψo表示捕获的激子将电子传递到QA以后的其他电子受体的概率,与对照相比,不同处理均降低叶片的RC/CSm值和Ψo值,与T1相比,T2、T3和T4处理分别降低了26.8%、68.4%、70.2%和11.2%、21.6%、40.8%。这表明臭氧胁迫下1600 μmol•m-2•s-1的强光和40℃高温均可加剧PSII最大光化学效率和潜在活性的降低,但1600 μmol•m-2•s-1的强光影响程度更大;当强光和高温同时出现时,抑制程度最大。
2.2 臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片Fv'/Fm'、ETR、qp的影响
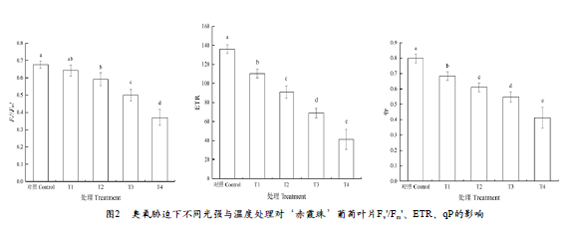
Fv'/Fm'是 PSⅡ有效光化学量子产量,为光条件下 PSⅡ天线转化效率,可以用来反映开放的PSII反应中心的激发能捕获效率,从图2可以看出,与对照相比,T1处理显著降低了植物叶片实际光化学效率,与T1相比,T2、T3分别降低了7.9%、22.1%,T4下降最显著,降幅为42.3%;ETR可以表示从PSⅡ到PSⅠ的电子传递情况[Munekageet al.],与对照相比,T1处理显著降低了植物叶片线性电子传递速率,与T1相比,T4处理降幅最大,达到62.5%,T2和T3处理分别降低了17.6%和37.5%;qP表示荧光淬灭中的光化学淬灭,不同处理显著降低了葡萄叶片的光化学淬灭,与T1相比,T2、T3 的qP值分别降低了10.7%和19.8%,T4变化最显著,降低了39.5%。这表明臭氧胁迫下1600 μmol•m-2•s-1的强光和40℃高温均可降低PSII激发能捕获效率、线性电子传递速率和光化学淬灭,导致叶片吸收的光能更多的以热耗散形式消耗,被用来进行碳固定的能量显著减少,但1600μmol•m-2•s-1的强光影响程度更显著;当强光和高温同时出现时,Fv'/Fm'、ETR和qP降幅最大,影响最显著。
2.3 臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片吸收光能分配的影响
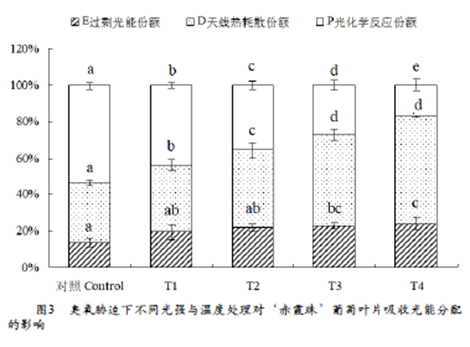
植物叶片吸收的光能有三条去路,一是在天线色素上进行热耗散(D),二是用于光化学反应(P),三是反应中心的过剩激发能(E)。图3反映了不同处理下吸收光能向光化学反应 、热耗散及过剩光能分配的情况,由图可知,与对照相比,不同处理下吸收的光能用于光化学反应的比例均有所下降,与T1相比,T2、T3分别降低18.8%和38.8%,T4下降最明显,降幅为62.0%。与T1相比,T2、T3和T4处理吸收的光能用于热耗散的比例均增加,增幅分别为15.9%、36.2%和60.5%;过剩光能分别增加12.5%、19.1%和25.2%。这表明臭氧胁迫下1600μmol•m-2•s-1的强光和40℃高温均可增加吸收的光能用于热耗散和过剩光能的份额,降低用于光化学反应的比例,但1600μmol•m-2•s-1的强光造成过剩光能的份额更大;当强光和高温同时出现时,过剩光能的比例最大;这部分既没有被捕光天线耗散掉,也没有被光化学反应利用的过剩光能必然会导致葡萄叶片活性氧含量增加,进而对光合机构造成损伤。
2.4 臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片光系统I、光系统II间激发能分配的影响
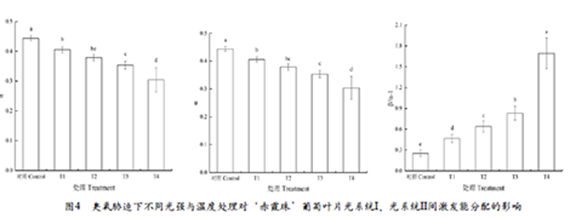
由图4可知,与对照相比,不同处理显著降低了叶片PSI激发能分配系数(α),与T1相比,T2、T3分别下降了6.6%和12.8%,T4下降幅度最大,降幅为25.1%;与之相反,不同处理后叶片PSII激发能分配系数(β)均显著上升,与T1相比,T2、T3、T4的分别升高了4.5%、8.7%、17.1%。双光系统间激发能分配不平衡偏离系数(β/α–1)变化趋势与PSII激发能分配系数一致,与T1相比,T2、T3分别升高37.7%和78.3%,T4升高幅度最大,达到187.8%。这表明臭氧胁迫下强光、高温严重影响激发能在两个系统之间的均匀分配,导致光系统之间激发能分配的不平衡,但强光的影响更为严重;当强光、高温同时出现时激发能在两个光系统间分配偏离最严重。
2.5 结论
40℃的高温、1600 μmol•m-2•s-1的强光均加剧了臭氧胁迫对PSII造成的伤害,但1600 μmol•m-2•s-1的强光影响程度显著大于40℃的高温;当强光和高温同时存在时,O3胁迫对PSII的伤害最明显,不仅加大了强光因素引起的光系统II伤害,也加大了高温因素引起的光系统II伤害。在葡萄大田栽培中,可以通过适当改变架势和叶幕降低光照强度来减缓臭氧伤害,通过喷施Ca2+缓解高温伤害也可以同时降低臭氧危害,减少强光和高温对臭氧危害的加剧作用。