鲜食葡萄栽培岗位
高世敏 郑焕 陶建敏
摘 要:葡萄果形的变化通常是由细胞分裂和细胞膨大共同协调控制的。赤霉素在果树生产上应用广泛,可以有效地保花保果,诱导无籽果实,也可促进果实膨大,提高果实品质。本研究中,外施GA3处理葡萄幼果可以调控葡萄果实形状。GA3可以使葡萄果实变长,果形指数显著增高。组织细胞学分析表明,GA3促进了葡萄果皮纵向细胞伸长。实时荧光PCR结果显示,赤霉素合成关键基因VvGA20ox2、VvGA3ox1基因在GA3处理后表达下调,而赤霉素钝化基因VvGA2ox1基因表达上调。GA3上调了细胞膨大伸长相关基因Vvexpansin-A8、VvXET23的表达,而抑制了细胞分裂有关基因VvCDKB2-1的表达。果形相关基因VvSUN基因在GA3处理后表达量升高,与对照相比,VvOFP1表达量则降低,这些结果表明,适宜的GA3浓度可以改变葡萄果实形状,并且影响相关基因的表达。
关键词:葡萄;赤霉素;果形
赤霉素作为一种重要的内源激素,在种子萌发、茎和根的伸长、叶片伸展、花和果实的发育等植物生长发育进程中起着重要的调控作用(Hedden P et al.,1997;黄桃鹏等,2005;Eriksson S et al.,2006;Hartweck et al.,2006)。果树生产中多用GA3来打破种子休眠,促进果树生长,促进节间变长(李洪明 等,2013)。在开花前用赤霉素处理葡萄花序,可拉长花序;在花期对葡萄进行赤霉素处理可使其无核化增高,提高坐果率;在花后对葡萄进行赤霉素处理可使果实增大,促进葡萄果实成熟(陶建敏 等,2003)。前人研究证明赤霉素对于增大果形指数有显著的作用(杨玉艳 等,2012;赵荣华 等,2014)。
葡萄是世界上广泛种植的果树之一,中国的鲜食葡萄产量占全球葡萄产量的 14%(Khalilurrehmanet al.,2017),金手指葡萄(GoldenFinger,V. vinifera L. × V. larbrusca L.)为鲜食葡萄的一种,甘甜爽口,有浓郁的冰糖味和牛奶味,且果粒形状奇特美观,深受广大消费者喜爱。由于自然环境及自身生长因素的影响,在成熟时葡萄果粒往往形态大小不统一,造成商品质量下降。为了更加有效地提高生产质量,展开葡萄果实形状的研究变得尤为重要。然而传统的改良果实形状的育种方法主要是杂交实生选种,这需要较长的时间和较大的种植规模(柴丹丹,2008)。
目前生产上多使用植物生长调节剂调节果实发育,改善果实品质,GA3作为一种重要的生长调节剂,在果实的发育及膨大方面起着重要作用,可调节果实形状(Davies et al.,2009; Wanget al.,2012)葡萄果实发育早期是葡萄形态建成极为重要的阶段,本试验通过外源GA3改变葡萄果实发育早期的内源赤霉素浓度,通过果实生理指标的变化及相关基因的表达来分析外源GA3对葡萄果实形状的影响。
1 材料与方法
1.1 材料
1.1.1 试验材料
试验于2017年在南京农业大学汤山葡萄试验基地进行,以 ‘金手指’葡萄为试验材料。采用平棚架避雨栽培,株行距为3.0m×6.0m,T树型。土肥水管理及病虫害防治同常规管理。以花序穗肩的第一个小穗上50%的小花开放为盛花期,于盛花后7d,随机选取树势相同的3株‘金手指’葡萄植株,使用25mg•L-1的赤霉素溶液对幼果进行浸泡处理,以蒸馏水浸泡作为对照,为增强试验的可靠性,分别在每棵树的一支臂进行处理,另一臂作为对照。每个处理1株;分别于处理后1d、3d、7d、14d随机采集处理及对照的小穗。采集的样品立即用液氮速冻,随后带回实验室,保存于-70℃冰箱中。
1.1.2 主要试剂
多糖多酚植物组织总RNA提取试剂盒购买于成都福际公司,rTaq酶,反转录试剂盒PrimeScript™ RTreagent Kit with gDNA Eraser和荧光染料SYBR Premix EX TaqTM购买于大连宝生有限公司(TaKaRa),RNA酶清除剂购自北京华越洋公司,有效成分为3%赤霉酸乳油购自山东鲁杭生物农药有限公司,96孔荧光定量PCR板购自于BIO-RAD公司。
1.1.3 主要仪器
冷冻离心机,电泳仪,凝胶成像系统,One DropTMOD-1000分光仪(Thermo Fisher Scienntific,USA),荧光定量PCR仪(ABI 7300system)、体式显微镜、载玻片、盖玻片、镊子、刀片等。
1.1.4 引物
利用Beacon Designer8软件设计荧光定量PCR引物(表2-1),荧光PCR引物由北京擎科新业物技术有限公司合成。
1.2 方法
1.2.1 GA3处理
选取3株长势一致的金手指葡萄树,为了增强试验的可靠性,在处理前统一修剪花序,序尖留约4cm,试验进行3次生物学重复,在每株树的两臂分别为处理组和对照组。以每个花序一半以上的小花开放时期作为盛花期,在盛花后一周,用25 mg•L-1 GA3溶液对幼果进行浸泡处理,处理时间大约为10s,以清水处理作为对照(CK)。
1.2.2 葡萄果实相关参数分析
在处理后1d、7d、15d、30d、45d、60d 6个时期采集果实样品,用冰袋带回实验室。每个处理随机选取20个果粒用游标卡尺测量其果实纵横径(每个组8个重复),统计数据,果实的纵径与横径的平均比值即为该果实的果形指数。
1.2.3 果皮细胞的细胞学观察
本研究选取处理后果形变化明显的时期的样品制作临时装片,每个样品采取3个重复。具体步骤如下:
(1)取洁净载玻片,于载玻片中央加上一滴水,将洁净盖玻片放好备用。
(2)用刀片用左手拇指、食品指和中指夹住新鲜葡萄幼果,样品材料,使其稍突出在手指之上,拇指略低于食指,材料和刀片蘸水,使其湿润。右手拇指和食指横向平握刀片,用刀片轻划出20-30mm长度的区域,用镊子将表皮撕下,迅速放入载玻片的水滴上,盖上盖玻片,轻敲盖玻片,使其平展。
(3)扫描电镜观察果实表皮细胞的变化并照相。
1.2.4 总RNA提取
采用成都福际公司的多糖多酚植物组织总RNA提取试剂盒提取RNA,具体操作步骤如下:
(1) 向2 m L 离心管中加入500μL Buffer PS1和10μLβ-巯基乙醇,混匀。
2) 取适量的葡萄组织材料,在液氮中迅速充分研磨成粉末,称取50mg粉末(葡萄果实可适当加量),快速转移至Buffer PS1中,在涡旋仪上剧烈震荡混匀,室温静置5min。
(3) 向上述液体中加入100μLBuffer PS,轻柔混匀。
(4) 将所有的液体转移至DNACleaningColumn中,13,300rpm离心2 min,去除过滤柱,并保留收集管内的滤液。
(5) 将滤液转移至新的2mL离心管中,加入1.5倍体积的BufferPS2,约800-900μL,轻柔混匀。
(6) 将混合液分两次转移至RNA-only Column中,每次不超过800 μL,12,000 rpm离心1 min,弃滤液。
(7) 向RNA-only Column中加入500 μL Buffer PRW1,12,000rpm离心1 min,弃滤液。
(8) 向RNA-only Column中加入700μL 无水乙醇,12,000 rpm离心1 min,弃滤液。
(9) 向RNA-umn中加入700μL Buffer PRW2,12,000 rpm离心1min,弃滤液,重复操作1次。
(10)将RNA-only Column放回收集管中,12,000rpm空管离心2min,弃掉收集管。
(11)将RNA-only Column放入新的1.5mL离心管中,加入30-50μL65℃预热的RNase-Free ddH2O, 室温静置2min,12,000rpm离心1min收集RNA溶液。
(12)将离心得到的RNA溶液重新加入RNA-only Column中,静置离心。取2μL RNA溶液进行琼脂糖凝胶电泳检测质量,取1 μLRNA溶液用分光仪测定浓度和纯度。
1.2.5 cDNA第一链的合成
c D N A 第一链的合成参照PrimeScript™ RT reagent Kit withgDNA Eraser操作说明书进行,具体步骤如下:
(1) 消化R N A 中的基因组DNA,在冰上配制消化混合液,10μL消化体系为:总RNA 1ug,5×gDNA Eraser Buffer 2μL,gDNA Eraser 1μL,用RNA FreeddH2O 补足10μ L ;42℃ 消化2min,4℃保存;
(2) 反转录合成cDNA第一链,在冰上预混反应液,混匀分装10μL到每个PCR管中,轻柔混匀,进行反转录反应。20μL反应液的体系为:消化后的RNA溶液10μL,PrimeScript RT EnzymeMixⅠ1μL,RT Primer Mix4μL,PrimeScript Buffer 2(for RealTime) 4μL,RNA Free ddH2O1μL。反应程序:37℃,15min;85℃,5 s;4℃保存。
1.2.5 实时荧光定量PCR
本试验采用实时荧光定量PCR方法测定各类基因在GA3响应后在葡萄幼果中的相对表达水平。
所用引物见表2-1,以葡萄 A c t i n (XM_002265440)作为内参基因。PCR采用20μl 反应体系:cDNA 1.2μl,2×SYBR PremixEX Taq TM (TaKaRa)10μl,上、下游引物各 0.4μl,灭菌后ddH2O补足至20μl;反应程序为:95℃4 min;95℃20s,60℃20 s,72℃40s,40个循环。每个样品设3个重复,采用ABI7300system 软件和 2 -ΔΔCT (K.J. Livak, et al.,2001)方法分析数据。


2 结果与分析
2.1 外源GA3对葡萄果实形状的形态学的影响
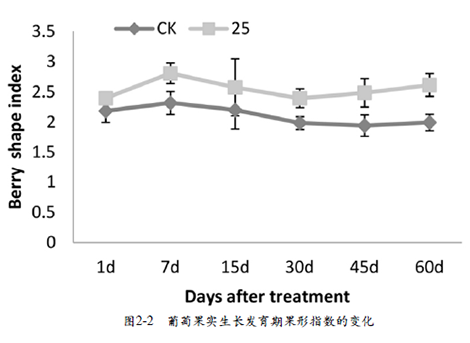
在本试验中,与对照相比,施用GA3后葡萄果实明显伸长(图2-1,D、E、F),果形发生显著改变,由‘弓形’变为‘长形’。为了更清楚地了解果形的变化程度,本试验采用果形指数来描述处理与对照果实在一段时期内果形的变化差异。结果表明,处理后葡萄幼果在GA3诱导后7 d果形指数达到最高(图2-2),说明在此时期果实的伸长最明显,随着果实的发育,果实开始进入发育的第二阶段,果实开始膨大,果形指数有所降低。在果实成熟时,处理组与对照组的果形指数达到第二个高峰,处理组的果形指数显著高于对照组的。以上结果说明,在金手指葡萄盛花后一周使用25mg•L-1 GA3处理葡萄幼果促进果实伸长,改变果形。
2.2 外源GA3对葡萄果实果皮细胞的影响
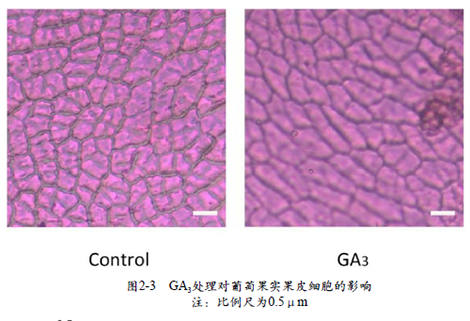
本试验以处理后7 d对照组与处理组的葡萄幼果为材料进行果皮细胞观察,通过比较同一部位的处理组与对照组果实果皮细胞来进一步研究GA3对葡萄果形的影响。结果显示,在同一视野下,GA3处理后的果皮细胞数量明显少于对照组的细胞数量,并且GA3处理组的果实表皮细胞在形态学上呈现拉长状(图2-3),说明GA3诱导果实变长可能与细胞伸长有关。
2.3 外源GA3处理对赤霉素合成相关基因表达的影响
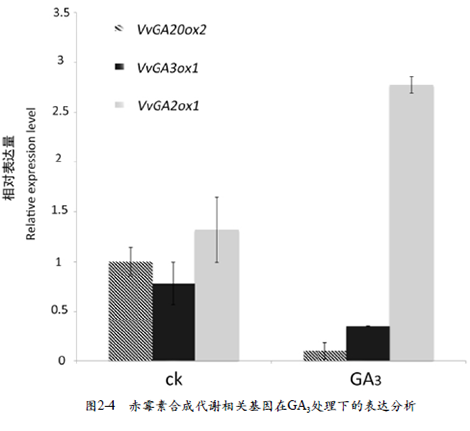
本试验以GA3处理后1 d的幼果提取RNA反转录成cDNA为模板,对赤霉素合成关键基因在果实中的表达进行分析。结果表明,GA3抑制了VvGA2 0ox 2、VvGA3 ox1基因的表达,促进了VvGA2ox1基因的表达(图2-4)。此结果与GA3响应番茄相关赤霉素合成代谢基因的表达结果一致(陈申,2016)。
2.4 外源GA3处理对细胞膨大和分裂相关基因表达的影响
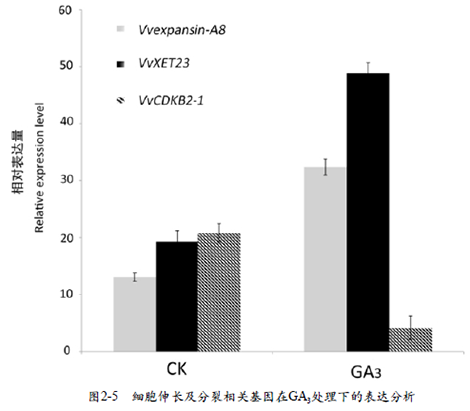
本试验选取与细胞膨大伸长有关基因expansin-A8、XET23及细胞分裂相关基因CDKB2-1在果实中的表达进行分析。结果显示,与对照相比,Vvexpansin-A8、VvXET23基因在GA3处理后1d上调表达,VvCDKB2-1 基因在处理后表达量下降( 图2-5)。以上结果说明,外源GA3可以调控果实发育前期有关细胞膨大与分裂的基因的表达,从而影响葡萄果形。
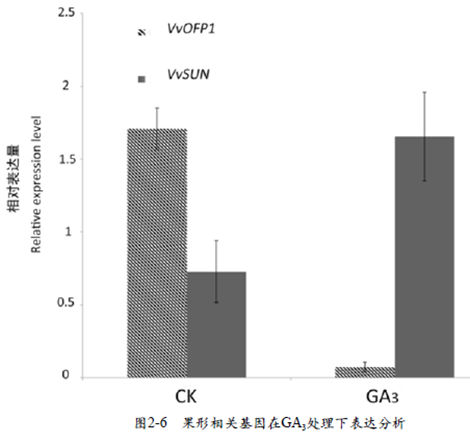
2.5 外源GA3处理对果形相关基因表达的影响
本试验通过对果形相关的基因OFP1、SUN基因在果实中的表达情况进行分析,以期从基因的表达水平变化上推测出外源GA3与果形相关基因之间的关系。结果表明,GA3处理后VvOFP1基因的表达量低于对照,而VvSUN基在GA3处理后上调作用明显(图2-6)。
3 讨论
关于赤霉素促进葡萄果实膨大方面的研究已有报道(姜卫兵,1992;吴俊,等,2001;谢周等,2010),由于外源激素在不同使用时间及浓度对葡萄果形的影响不同,且根据葡萄品种的特异性,难以掌握准确的方式有效地改变果实形状,通过分子生物学方法研究葡萄果形变化的机理变得尤为重要,有助于今后的生产及理论研究。本试验以25 mg•L-1 GA3处理金手指葡萄盛花后7d幼果,发现处理过后的果实明显伸长,果形由弓形变为细长形,从表型上的改变证实了GA3可促进葡萄果实伸长的作用。本试验以处理后7 d对照组与处理组的葡萄幼果为材料进行果皮细胞观察,通过比较同一部位的处理组与对照组果实果皮细胞来进一步研究GA3对葡萄果实的影响。结果显示,在同一视野下,GA3处理后的果皮细胞数量明显少于对照组的细胞数量,并且GA3处理组的果实表皮细胞在形态学上呈现拉长状,说明GA3诱导果实变长可能是由于细胞伸长导致。
本试验以GA3处理后1d的幼果提取RNA反转录成cDNA为模板,对赤霉素合成关键基因在果实中的表达进行分析。结果表明,GA3抑制了VvGA20ox2、VvGA3ox1基因的表达,促进了VvGA2ox1基因的表达。此结果与GA3响应番茄相关赤霉素合成代谢基因的表达结果一致(陈申,2016)。
本试验选取与细胞膨大伸长有关基因expansin-A8、XET23及细胞分裂相关基因CDKB2-1在果实中的表达进行分析。结果显示,与对照相比,V ve x pa n sin- A8、Vv X E T23 基因在GA3处理后1 d上调表达, VvCDKB2 -1基因在处理后表达量下降。以上结果说明,外源GA3可以调控果实发育前期有关细胞膨大与分裂的基因的表达,从而影响葡萄果形。本试验通过对果形相关的基因OFP1、SUN基因在果实中的表达情况进行分析,以期从基因的表达水平变化上推测出外源GA3与果形相关基因之间的关系。结果表明,GA3处理后VvOFP1基因的表达量低于对照,而Vv SUN基在GA3处理后上调作用明显。
赤霉素具有促进细胞膨大伸长的作用已经得到大部分研究的证实(De Jong et al.,2009)。本试验为了从细胞学水平说明外源GA3与果实伸长之间的关系,以处理组果形指数最高时期的幼果为材料,进行果实表皮细胞观察,结果显示GA3促进了果皮细胞延长。从细胞学角度来看,果实发育前期的细胞分裂可能己经确定了其果形指数(Robinson MC Goffinet TL, etal.,1995)。
外源GA3促进果形伸长可能是由于内源活性赤霉素的改变引起,通过调控相关基因的表达变化来影响果实形状。植物内源活性赤霉素主要通过赤霉素代谢反馈调节机制来维持一定的平衡水平(Hedden et al.,2000;Ribeiro etal.,2012)。本试验中,GA3促使赤霉素合成基因基因Vv G A 3 o x 1、VvGA2 0ox 2基因下调,而赤霉素钝化基因VvGA2ox1基因表达上调。葡萄生长由细胞膨大和细胞分裂完成,细胞分裂与一些细胞周期蛋白相关,由研究表明,CDKB2是在细胞分裂阶段高表达的一个重要的细胞周期控制基因(Czerednik, A. etal.2012)。expansin基因在调节细胞可塑性方面有着重要作用,与细胞大小及形状的变化有关(FukudaH et al.,2014)。木葡聚糖内糖基转移酶蛋白(XET)主要在细胞壁松弛作用起作用(L.G. Delucet al.,2007),其主要机理是与expansin基因协同促进交联聚糖微纤维松动,造成其分离,改变细胞渗透压,导致细胞膨大(Buchananet al., 2000),在百合的花瓣发育过程中XET基因上调表达(Laitinenet al. 2007)。
本试验在GA3处理后VvCDKB2-1基因表现为抑制,而Vvexpansin-A8、VvXET23基因表现为上调。说明GA3诱导葡萄果实形状可能与细胞膨大和分裂基因存在一定的关系。
在之前的报道中,番茄OVATE 基因的超表达,导致植株的多个器官生长受到抑制,表现为植株矮小,花冠、花丝等花器官变短,小叶变圆,柱头外露等 (Liu etal.,2002)。OsOFP2和BLH-KNOX复合物相互作用抑制木质素的合成,以及OsGA20ox7基因表达,调控水稻的生长发育(Schmitz et al.,2015)。
辣椒C a O VAT E 蛋白也是通过抑制CaGA20ox1表达,从而改变果实形状(Tsaballa et al.,2011)。 SUN 基因编码了一个 IQ67 结构域的植物蛋白成员,为 IQD12,该蛋白可以独自实现使番茄表现长果形而非圆果形的变化(Han, et al., 2008)。SUN 基因在长果形番茄中高表达,而在圆果番茄中不表达,在圆果番茄中转入 SUN基因则长出狭长的果实,再将长果实的SUN基因敲除后则出现圆形果(van der Knaap et al.,2004)。本试验中GA3促使VvOFP1 基因下调表达,而VvSUN基因表达上调,与之前研究结果一致。综上,VvOFP1、VvSUN基因可能参与了葡萄果实发育过程中形状的调控。而葡萄果形调控是一个复杂的过程,仍需通过转录组测序、基因功能验证等方法进一步进行研究。