鲜食品种改良岗位
类黄酮化合物是一大类多酚类次生代谢产物,对葡萄及葡萄酒色泽、风味、收敛性及苦味等重要品质的形成具有重要作用(Flamini etal.,2013),同时具有抗氧化、抗癌、抗衰老、保护心血管等药理学作用(Georgiev et al.,2014)。类黄酮的合成和调控目前是生命科学研究的热点之一。
查耳酮异构酶( C H I ; E C5.5.1.6)是类黄酮代谢途径的入口酶,也是此途径的一个关键限制酶,催化4, 2′, 4′, 6′ -四羟基查耳酮、4, 2′, 4′ -三羟基查耳酮、2′, 4′ -二羟基查耳酮以及4, 2′ -二羟基查耳酮生成相应的类黄酮物质(Jez et al.,2000),使双环的查耳酮变成了三环的类黄酮类物质的骨架结构。在葡萄中,CHI基因几乎在所有的葡萄组织中(果实、叶片、茎、根等)都有表达(Bosset al.,1996)。但是由于CHI和其他类黄酮合成酶一样,在葡萄中的浓度很低很难去检测(Robinson& Davies,2000),所以多数关于CHI的信息主要来自于基因水平的研究。而对于基因表达的研究并不能反映蛋白的积累情况,明白其蛋白的表达规律及在细胞中的定位对于深入理解类黄酮合成及调控的细胞学机制具有重要的意义。
关于C H I 的亚细胞定位,Saslowsky和Winkel-shirley(2001)采用免疫电子显微技术和免疫荧光技术在拟南芥幼苗根部对CHI进行了亚细胞分布观察,结果表明CHI分布在内质网和液泡膜上(Saslowsky & Winkel-shirley.,2001),2005年又在拟南芥根尖伸长区表皮细胞细胞核中检测到了CHI的分布(Saslowsky et al.,2005)。然而这些研究都主要集中于CHI在营养器官中的亚细胞定位,关于其在生殖器官,例如果实中的亚细胞分布还尚不清楚。
之前的研究已经克隆了葡萄CHI基因全长并且制备了CHI多克隆抗体(Wang et al.,2012),本研究中利用该抗体,采用蛋白免疫杂交及胶体金免疫技术对葡萄果实发育过程中CHI表达及亚细胞定位进行了分析,为深入了解葡萄果实中不同分支类黄酮产物的合成和调节提供新的见解和思路。
1 材料与方法
1.1 材料
以北京龙徽酿酒有限公司河北怀来县葡萄生产基地1997年定植的酿酒葡萄品种‘
赤霞珠’(Vitis vinifera L. ‘Cabernetsauvignon’)果实为试材,选择结果枝粗细一致,结果部位一致的果实。2014年从花后20 d开始取样,每10 d取一次,一直取样到葡萄浆果完全成熟(约花后120 d),每次随机取果实50穗,去除小次果粒。立即进行固定处理或者液氮速冻保存于-80°C冰箱,进行后续试验。葡萄C H I 多克隆抗体为本实验室自己制备(Wang et al.,2012)。
1.2 葡萄果实总类黄酮、RNA提取及CHI基因表达分析
葡萄果实总类黄酮含量的提取测定参照Wang等(2012)的方法。
葡萄果实RNA提取及cDNA制备参考Wang等(2010)的方法。根据 GenBank 上发表的葡萄CHI(X75936)和Actin1(AY680701)序列设计PCR扩增引物(表1),引物由生工(上海)股份有限公司合成。目的基因的扩增参考Wang等(2012)的方法。
1.3 葡萄果实蛋白提取及蛋白质印记杂交
葡萄果实蛋白质的提取参考Famiani等(2000)的方法稍做修改。精确称取经液氮研磨后的果实1g,加入2mL 提取液(50mmol·L-1Tris-HCl,(pH8.9),2% SDS,5mmol·L-1 抗坏血酸,5m mol·L-1EDTA,14m mol·L-1 β-巯基乙醇,10μmol·L-1 Leupeptine,1m mol·L-1PMSF,0.15% PVP)。样品充分混匀后进行超声波破碎3min,4层纱布过滤后,4 ℃,12000×g 离心15min,取上清液即为可溶性蛋白质提取液。蛋白含量的测定参考Bradford(1976)的方法,以牛血清蛋白(BSA)为标准。蛋白质免疫印记杂交分析参考Wang等(2010)的方法。
1.4 胶体金免疫定位
参照Chen 等(2006)的方法,将葡萄果皮切分成2~3 mm3的小块,迅速放入已预冷的4%多聚甲醛与2.5%戊二醛(100 mmol·L-1的磷酸缓冲液配制,pH 7.2)的混合固定液中4℃下固定4 h。经过预冷的磷酸缓冲液在4℃下洗涤3次,每次30min,之后乙醇系列脱水,然后采用乙醇-Lowicryl K4M 置换及纯K4M 浸透过夜,-20℃下紫外聚合48h,室温继续聚合48h。用LKB-8800 型切片机制备超薄切片,将切片置于100目以0.3%Formvar膜覆盖的镍网上。
参照Wa n g 等( 2 0 11 )
的方法, 切片经过前处理后, 在T B S T + 0
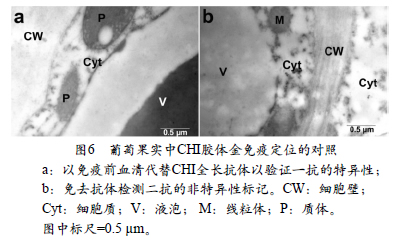
. 1 gL-1BSA 缓冲液稀释5 0 倍的一抗(CHI全长抗体)中37 ℃下温育处理3h。之后用TBST+0.2g L-1BSA 室温下冲洗3次,每次5 min,然后转入羊抗兔金标记二抗中(10 nm)(1:50)37 ℃温育处理60 min。充分洗涤后用2%的醋酸双氧铀25 ℃下染色20min,重蒸水冲洗6次,每次5min,柠檬酸铅25 ℃下染色15 min,用重蒸水冲洗6次,每次5 min,透射电镜观察。为了检验免疫电镜定位结果的特异性和可靠性,分别设置了两种不同的对照:(1)以免疫前血清代替全长抗体以验证一抗的特异性;(2)免去抗体以检测二抗的非特异性标记。其余步骤同上。多次重复。
2 结果与分析
2.1 葡萄果实发育过程中总类黄酮含量的变化

从图1可以看出,总类黄酮含量在葡萄果实发育早期较高,之后逐渐降低,到转色期又快速升高。在80 d和90 d天略有降低,到成熟采收时
达到最大值。
2.2 葡萄果实发育过程中CHI基因和蛋白表达的变化
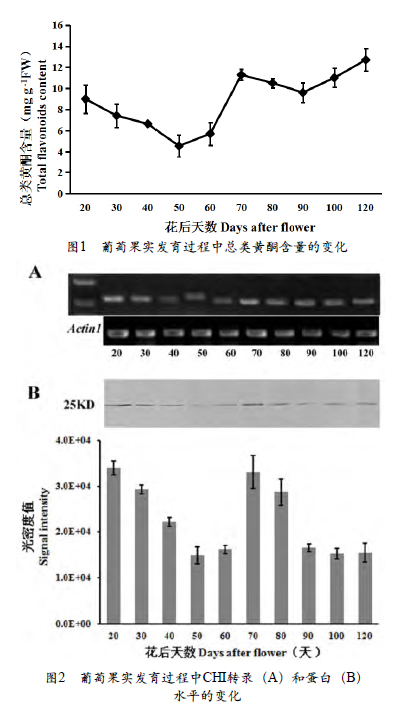
如图2,A所示,CHI基因在果实发育早期(花后20 d)转录处于很高的水平,随后急剧下降,至转色期(花后70 d)又迅速上升,之后一直保持较高的水平。利用葡萄CHI抗体,从葡萄果实总蛋白中检测到一条蛋白条带(大约25 KD,图2,B),确定为目的蛋白。在葡萄果实发育过程中,CHI蛋白的积累形式与基因水平相似,也是在果实发育早期(花后20 d)积累最多,在缓慢生长期逐渐下降,紧接着在转色期又迅速积累,在第2次快速生长后期出现第2次下降直至花后100 d,在成熟采收时(花后120 d)又有所升高。
2.3 葡萄果实发育过程中CHI亚细胞定位
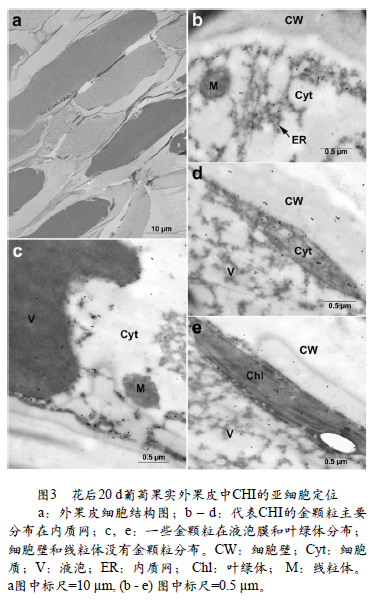
大量研究表明类黄酮类物质主要在葡萄果实果皮中合成积累(Braidot et al.,2008),本文着重研究了葡萄果实发育过程中果皮细胞中CHI的定位。图3显示葡萄果实发育早期(花后20 d)果皮细胞中CHI的分布。图3,a表明细胞具有完整的细胞结构。代表CHI蛋白的金颗粒主要分布在细胞质的内质网上(图3,b,d),少量金颗粒在液泡(图3,c,e)和质体(主要是叶绿体)也有分布(图3,e)。而在线粒体和细胞壁没有发现金颗粒的分布。
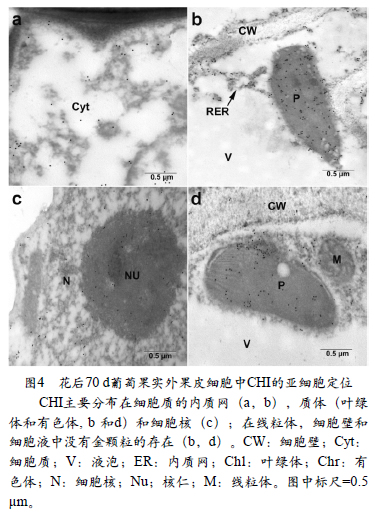
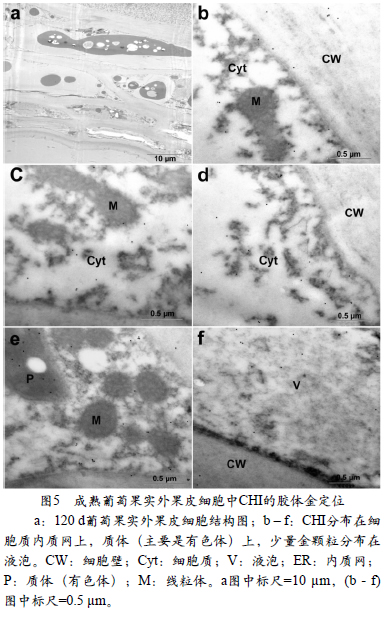
图4显示了CHI在转色期(花后70 d)果实外果皮细胞中的亚细胞分布。在转色期,花色苷开始合成并积累,一部分叶绿体转变为有色体(图4,b和d)。代表CHI的金颗粒主要分布在细胞质的内质网上(图4,a,b)和质体(叶绿体和有色体,图4,b和d)中,此外在细胞核中也发现了CHI的大量分布(图4,c)。CHI的数量在此时期显著增加。在成熟葡萄(花后120 d)果实外果皮细胞中(图5),代表CHI的金颗粒仍然主要分布在细胞质(图5,b ~ e),在质体(有色体)(图5,d,f)和液泡中也有少量金颗粒的分布(图5,c)。在线粒体和细胞壁上面没有金颗粒的分布。在这个发育阶段,相较于转色期, CHI明显减少。
本试验设立了2组对照用来检测CHI免疫电镜定位结果的特异性和可靠性。如图6,a,b所示,两组对照中都没有金颗粒的存在,体现了试验方法和结果的可靠性。
3 讨论
本研究中,分别在蛋白和转录水平观察了CHI的表达变化。在葡萄果实发育早期,CHI呈现高水平的表达,这时类黄酮大量积累。接着CHI表达下降,类黄酮的积累也有所下降。随着果实转色期的到来,CHI表达水平再次升高,伴随着类黄酮第二次积累高峰的到来,这与前人的研究结果一致(Wanget al.,2012)。其他类黄酮合成基因的表达水平也相应提高(Wang etal.,2010;Wang et al.,2016)。这表明在葡萄果实发育过程中,CHI受到蛋白水平和转录水平双重调控,并且CHI的调控具有时空特性。
对葡萄果实中类黄酮代谢相关酶进行亚细胞定位的研究,对于了解果实次生物质代谢的细胞学机制具有重要意义。许多证据表明类黄酮代谢途径中的酶以多酶复合体的形式存在。在细胞中,类黄酮合成相关酶围绕着细胞色素P450酶类共定位于内质网上(Winkel-shirley,2004)。然而,之后的研究发现类黄酮合成相关酶存在更复杂的定位模式,不仅仅定位于细胞质中的内质网,可能还定位于液泡、质体及细胞核等细胞器中(Petrussaet al.,2013)。CHI是类黄酮代谢的一个关键入口酶,其在葡萄果实果皮细胞中的细胞学定位尚未见报道。本研究中在葡萄果实不同发育阶段(幼果期、转色期、成熟期)对该酶进行了胶体金免疫定位。
研究结果表明在整个果实发育过程中CHI在细胞质的内质网上都有分布。同时在液泡中也发现有CHI的少量分布。这与之前Saslowsky和Winkel-shirley(2001)的结果一致。CHI的数量在不同的发育时期有所变化,在果实发育早期有一定积累,到转色期显著增加,果实成熟期又有所下降,这与CHI蛋白在果实发育过程的积累模式相似。转色期葡萄果皮细胞中CHI数量的显著增加可能与这个时期类黄酮类物质大量合成有关。
本研究中还发现,在不同发育时期葡萄果实果皮细胞中,CHI在质体(叶绿体和有色体)中也有一定的分布。
尽管这种质体定位模式很少报道,但是之前的研究发现葡萄果实肉桂酸-4-羟化酶(C4H)、查尔酮合成酶(CHS)和花青素合成酶(ANS)也分布在外果皮细胞的质体中(Chen et al.,2006;Tian etal.,2008;Wang et al.,2010),表明在葡萄外果皮细胞质体中还有其他类黄酮合成酶的分布,这些酶可能连同CHI组成多酶复合体形式以利于类黄酮的合成(Winkelshirley,2004)。在转色期葡萄果实果皮细胞中,CHI在细胞核也有分布。Saslowsky等(2005)在拟南芥根尖细胞中也发现CHI定位在细胞核中。在葡萄果实原生质体中CHI也表现出细胞核定位模式(Wang et al.,2015)。同时在许多植物的细胞核上也发现了类黄酮类物质的积累(Saslowsky et al.,2005;Polster et al.,2006)。这些结果表明可能有类黄酮类物质在细胞核原位合成,但是这还有待进一步验证。
总之,本研究发现在葡萄果实发育过程中,CHI表现出细胞质(内质网)、质体及细胞核定位模式,这样的定位模式可能由葡萄果实中类黄酮代谢的丰富和多分支造成的。继续探究造成不同定位的原因对于未来调控和改变葡萄果实的次生代谢具有重要的意义。