鲜食葡萄栽培岗位
侯旭东
陶建敏
摘 要:【目的】探究不同浓度GA3(赤霉素)和CPPU[N-(2-氯-4-吡啶基)-N'-苯基脲)]处理对‘阳光玫瑰’葡萄果锈的形成机理。【方法】在盛花两周后分别用25mg·L-1GA3 (A),25mg·L-1GA3+5 mg·L-1CPPU(B),25mg·L-1GA3+10mg·L-1CPPU(C),25mg·L-1GA3+15mg·L-1CPPU(D)对葡萄幼果进行处理,清水处理作为CK。利用酶标仪测定不同处理在果锈出现的时期(花后14周)木质素的含量,以及盛花后9、11、13、15和17周果皮中各处理相关酶活性变化;利用试剂盒提取果锈形成关键时期(花后13周)CK、处理A和处理C的RNA,然后进行RNA-Seq测序,并对其结果进行分析,在苯丙烷合成代谢途径中筛选出6个与酶活性相关的差异基因,利用RT-qPCR测定不同处理的相对表达量,并对其结果进行验证。最终把不同处理间木质素、酶活性和基因表达的变化趋势进行比较分析。【结果】 CK表面的果锈分布最多,其次是处理A,处理B和处理D表面的果锈较少,处理C表面光滑,色泽鲜艳,基本没有果锈的出现。CK的木质素含量显著高于其他处理,处理25mg·L-1GA3+10mg·L-1CPPU(C)的木质素含量最低。PAL酶活性的变化呈先下降后上升的趋势,花后13周,酶活性由高到低依次是CK、D、A、B、C;在花后第9周CK与各处理间C4H酶活性的差异不大,第11周后有所上升;不同处理对4CL酶活性的影响呈“W”的变化趋势,第13周4CL活性为CK>处理A>处理D>处理B>处理C;不同浓度植物生长调节剂处理对POD酶活性的影响呈先下降后持平的趋势;不同浓度植物生长调节剂处理对PPO酶活性的影响整体呈“V型”的趋势。RTqPCR结果与RNA-seq测序结果基本一致,在筛选出的基因中POD (LOC100242338)和PAL (LOC100233012)的表达量与木质素含量变化趋势基本一致,CK的表达量显著高于处理C。【结论】25mg·L-1GA3能减少果锈的发生,在此基础上10mg·L-1CPPU能基本清除果锈的发生。即处理C可能能够通过抑制LOC100233012的表达,来降低PAL活性,从而抑制木质素的合成来减弱木栓化,最终达到减少葡萄果锈来提高其商品价值的目的。另外,4CL酶的升高也可能在果锈减少的过程中起到作用。
关键词:GA3;CPPU;‘阳光玫瑰’葡萄;果锈;木质素;酶活性
‘阳光玫瑰’葡萄属欧美杂交种,是日本果树研究所用‘安芸津21号’和‘白南’为亲本杂交育成的新品种,二倍体葡萄品种。该品种外形美观,果肉酸甜适口,脆而多汁,具有浓郁的玫瑰香味。可溶性固形物含量达20%~23%,最高可达28% ,具有晚熟,挂果期长等特点。由于‘阳光玫瑰’抗病性强,花芽分化好,易于栽培,近些年在国内的种植面积在逐渐的增加。但阳光玫瑰在自然生长条件下果粒偏小,果穗稀疏并且易出现果锈,严重的影响了其商品价值。
赤霉素(gibberellin A3,GA3)和N-(2-氯-4-吡啶基)-N'-苯基脲[N-(2-chloro-4-pyridyl)-N'-phenylurea,CPPU]是作为植物生长调节剂广泛用于葡萄的生产栽培。GA3能打破休眠,促进种子的萌发、茎叶的生长、细胞的伸长、起到增大果粒的作用,还能起到预防落花落果的作用。CPPU具有促进细胞分裂和增大,延缓衰老等作用,因而能够增加产量,提高葡萄的商品性。
果锈又称水锈,是一种生理病害,表现为果面浮生的一层黄褐或赤褐色木栓化组织。果锈发生时会在果面形成黄褐色斑点,严重的影响了果实的商品价值。果锈的发生易受环境因素的影响,根据试验认为高湿、低温、通风不良以及光照不足等环境因素都会诱导果锈的产生。另外机械损伤也是诱发果锈发生的一个重要因素。通过喷施赤霉素、二氧化硅、杀菌剂以及套袋等方法可避免或降低果锈发生率,其中喷施赤霉素和套袋效果最为显著。
目前关于果锈发生的内在机理研究有很多,但主要有两种。国内学者吴厚玖等和李壮等认为果锈是从内向外产生的,即先是表皮和下皮的部分细胞接受某中刺激变成黄褐色,细胞发生异常分裂增殖形成木栓形成层,继而进一步分裂形成的木栓组织顶破角质层,黄褐色的果锈就是破损的角质层;国外学者SIMONS和LOONEY认为果锈是从外向内形成的,首先果皮受到外界环境的刺激后,角质层发生龟裂,外表皮下的细胞失去角质层的保护,直接与外界环境接触,受到刺激导致分裂增殖,促进木栓组织的形成,木栓化的周皮替代表皮所形成的黄褐色的物质就是果锈。但是两种机理在果锈形成的过程中都涉及到了“木栓化”这个共同的关键点。李国刚等认为较高的温度和湿度会促进果锈的形成,同时果皮的木质素和纤维素的含量比较高;还证明了出现锈斑区域的果皮木质素和纤维素的含量要高于完好果皮组织,而且果锈面积越大,木质素含量越高。
木质素的合成是个极其复杂的过程,大致可以分为三个阶段,分别是莽草酸代谢途径、苯丙烷代谢途径和木质素合成的特异途径。朱玲等认为苯丙烷代谢途径和木质素合成的特异途径为木质素合成的主要途径。苯丙氨酸解氨酶(PAL)是木质素单体合成过程中是第一个关键酶,也是苯丙烷代谢途径的第一限速酶。Kawaoka等认为抑制烟草中的Ntliml的表达引起了PAL、CAD和4CL基因表达水平的下降,从而导致木质素含量下降约27%。肉桂酸-4-羟基化酶(C4H)是苯丙烷代谢途径的第二个关键酶,当C4H酶活受到抑制时,植物体内的木质素含量明显减少。4-香豆酸辅酶A连接酶(4CL)是苯丙烷代谢途径的最后一个关键酶,在杨树中提高S木质素合成相关酶(PAL、C4H、4CL)的活性,会导致杨树愈伤组织中S木质素的含量上升。肉桂醇脱氢酶(CAD)在木质素单体合成的最后一步起关键作用。
有研究表明下调柳枝稷(Panicumvirgatum)中CAD的表达,植物中木质素的含量有所下降。过氧化物酶(POD)是催化木质素单体脱氢聚合最终形成木质素的酶,在木质素合成的特异途径中起关键作用。张华云等认为多酚氧化酶(PPO)和POD活性的降低会导致木质素的合成量减少。有研究证明,PAL、PPO和POD酶活活性的降低,会导致表皮细胞分泌蜡质减少,木质素合成减少,木栓形成层的发生和活化受到抑制。
本实验就是以‘阳光玫瑰’葡萄为试材,通过喷施不同浓度的GA3和CPPU,测定不同处理在不同时期的木质素含量和酶活活性的变化,分析木质素与酶活变化与果锈形成的相关性。对果锈形成关键时期的果皮进行转录组测序,然后在测序结果基础上,在苯丙烷合成代谢途径中筛选表达具有差异的基因。把筛选出来的基因进行设计引物,利用SYBR Green实时荧光定量PCR手段进行验证这些基因在不同处理间果皮中的表达差异。最后,综合对比分析不同处理间木质素、酶活性和基因表达的变化趋势,来进一步探究果皮锈斑形成的原理。
1 材料和方法
1.1 材料
供试材料是‘阳光玫瑰’葡萄不同发育时期的果皮,样品于2016年7-10月在南京农业大学汤山翠谷葡萄实验基地采取。采用平棚架‘H’型树形,进行避雨栽培,株行距为4.0 m×8.0 m,土肥水、花果管理及病虫害防治同常规。选取树势相似的葡萄树,在花后两周用不同浓度的植物生长调节剂浸蘸花穗5-10s,供试浓度为:25mg·L-1GA3(A),25mg·L-1GA3+5 mg · L - 1CPPU( B )
,25mg · L - 1GA3+10mg · L - 1CPPU(C),25mg·L-1GA3+15mg·L-1CPPU(D),清水处理作为CK。
1.2 方法
1.2.1 样品采集
从盛花期9周(9 DAW)开始样品采集,样品的采集注重随机性,每穗上中下随机采样,每株作为一个重复,共三个重复。每次采大小均匀、成熟度一致20粒左右,采样后立即放入冰盒,带回实验室剥下果皮,在超低温冰箱中进行保存。
1.2.2 木质素的测定
参照Hatfield R D(38)等,将花后14周的CK和不同处理的葡萄果皮用锡箔纸包裹,然后放到80℃烘箱烘干,研成粉末,用电子天平准确称取果皮0.02g各三份。将称取的样品粉末放入试管中,加入2mL25%的溴乙酰-乙酸溶液(w/w)和0.08mL高氯酸,封口后,70℃恒温水浴30min,每隔10min振荡试管。取四分之一的反应液,移入已装有1mL 2mol·L-1NaOH和2.5ml冰乙酸混合液的容量瓶内,充分振荡,用冰乙酸定容至10mL。以冰乙酸为空白溶液,用酶标仪对样品溶液在波长260nm处进行扫描测定吸光度。Lignin=Abs×liters×100%/W sample,其中Lignin为木质素含量,Abs为样品溶液木质素吸光度,Liters为样品溶液定容体积(mL),W sample为样品绝干重量(g)。
1.2.3 酶活性的测定
PAL活性测定参照Assis等的方法并稍作了些修改。称取0.2g果皮,放入预冷的研钵中,加入2mL0.1mol·L-1硼酸缓冲液(pH8.8),充分研磨后离心,
得2mL粗酶液。酶活力的测定体系包括:1mL粗酶液+2mL硼酸缓冲液+1mL0.02mol·L-1苯丙氨酸,调零对照为酶液换成硼酸缓冲液。混匀,30℃水浴1h,用酶标仪在290nm处测其OD值。
C4H活性测定参照Lamb等的方法稍作修改进行。提取液为2mL50mM,pH=8.9 Tris-Hcl(15mM巯基乙醇,4mM Mgcl2,2.5mM维C,10μM亮抑酶肽,0.15%聚乙烯吡咯烷酮,10%甘油),反应液为50mM,pH8.9 Tris-Hcl(2μM反式肉桂酸,2μM氧化型辅酶钠盐,5μM葡萄糖六磷酸二钠)。混匀后,25℃震荡30min,加入100μL6M盐酸终止反应,
摇匀, 4 ℃,5000g离心5min去除变性蛋白,在340nm处测其OD值。
4CL活性测定参照毕咏梅和欧阳光察等的方法稍作修改进行。提取液为2mL 0.2M,pH8.0 Tris-Hcl(含25%甘油,0.1mM DTT),反应液为0.2M,pH8.0
Tris-Hcl(15μMMgCl2 5μM P-香豆酸 50μM ATP1μM CoA)。混匀后,40℃水浴10min,加入100μL 6M盐酸终止反应,摇匀,4℃ 5000g离心5min去除变性蛋白,在333nm处测其OD值。
POD活性测定参照Briecka和李合生等的方法稍作修改进行。提取液为2mL 0.2M, pH6.0的磷酸缓冲液,反应液为0.2M,pH6.0的磷酸缓冲液(50mL含28μL愈创木酚,19μL 30%的H2O2)。混匀后静止10min,在470nm处测其OD值。
PPO活性测定参照罗晓芳等的方法稍作修改进行。提取液为2mLpH6.0的0.2M磷酸氢二钠-0.1M柠檬酸缓冲液,反应液为pH=6.0的0.2M磷酸氢二钠-0.1M柠檬酸缓冲液(内含0.05M邻苯二酚)。混匀后,30℃水浴10min,放入沸水中终止反应,在410nm处测其OD值。
所有酶活力测定样品为0.2g,酶活力的测定体系为:1mL粗酶液+2mL反应液,粗酶液换成提取液为调零对照。酶活力计算公式为:
U 酶活力= ( △ A · V t ) /(W·Vs·t·0.01)
其中:
△A为吸光度变化;W为样品重量( g )
; t 为反应时间(min);Vt为酶液总体积(mL);Vs测定所用酶液体积(mL)
。设置每分钟OD值变化0.01为一个酶活单位,单位u/(g·min)。
1.2.4 RNA-seq与实时荧光定量RT-PCR分析
用RNA提取试剂盒 (Foregene,Chengdu, China)按照说明书来进行样品总RNA的提取,所有RNA样品在1%的琼脂凝胶上进行检测,当A260与A280的比值大于1.8时,总RNA样品才能被检测出来,达到可以使用的纯度。总RNA样品被汇集到三个样品阶段,每个样品中10μg的总样品RNA会被用于建立Illumina RNA-Seq基因库,按照人工指令通过Illumina HiSeqTM2000进行测序。
以总R N A 为模板利用PrimeScriptTMRT-PCR(Takara)试剂盒反转录合成cDNA。内参基因为UBQ。结合RNA-seq测序结果,在NCBI上找出苯丙烷代谢途径中的关键酶基因,qRT-PCR引物用Beacon Designer 7(PREMIERBiosoft 公司)设计。引物由擎科生物工程有限公司(南京)合成。20μL PCR反应体系为:TakaraSYBR Premix Ex Taq(TaKaRa)10μL,上游引物和下游引物各0.5μL,稀释至200 ug·ml-1的 cDNA1μL和8μL去离子水。反应程序如下:95℃预变性4 min,95℃变性20s,60℃退火20 s,72℃延伸30 s,40个循环。
1.3 数据处理
木质素含量,酶活性的数据测定生物学重复3次,qRT-PCR经UBQ内参检验后生物学重复3次,所有数据处理通过SPSS Statistics17.0 软件处理(Duncan's和LSD检验,P<0.05)。
2 结果
2.1 同一时期不同处理果锈的分布
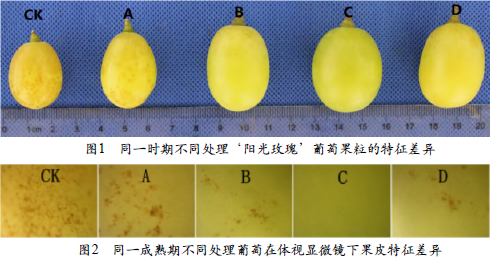
图1和图2为花后第14周不同处理葡萄果皮表面果锈的发生状况,显而易见,不同处理果皮表面果锈的附着程度存在明显的差异。CK果皮表面的果锈分布最为密集,也最为广泛,所占果皮表面积的比例也最高;处理A就较为次之,分布相对CK较为稀疏,但在果皮的各个部位也都有出现;处理B和处理D表面的果锈较少,只有在果实的果梗周围出现,附着的密度也非常稀疏;处理C表面的果锈在所有处理中最少,基本没有出现果锈,表面光滑,色泽鲜艳。
2.2 木质素含量的变化
图3是花后14周不同处理木质素含量的差异,CK木质素含量最高,处理A次之,处理C的含量最低,处理B和处理D的含量无明显差异,但略高于处理C。
2.3 相关酶活性的变化
2.3.1 PAL酶活性的变化
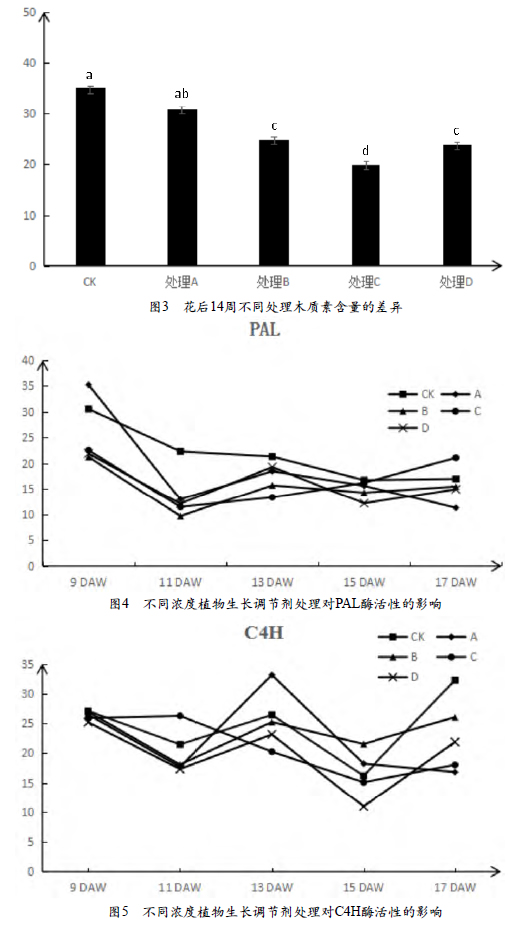
由图4能看出不同浓度植物生长调节剂处理对PAL酶活性的影响呈先下降后上升的趋势。CK的酶活性大部分时期都处在最高值,处理B和处理C的酶活性基本都比较低,处在折线图的最下方。花后第9周CK和处理A的值较大,处理B、C和D的酶活性基本相同;花后11周,所有处理和CK的值都呈现下降的趋势;花后13周,酶活性由高到低依次是CK、D、A、B、C;花后15周以后CK与各处理的酶活性都逐渐的减少了差距。
2.3.2 C4H酶活性的变化
由图5能看出,在花后第9周CK与各处理间C4H酶活性的差异不大;到第11周后有所上升,CK的酶活性较高,处理A、B和D的酶活性较低;在13周出现了短暂上升,酶活性由高到低的顺序分别是处理A、CK、处理B、处理D、处理C;在第15周各处理的酶活性都出现下降,酶活性变成:处理B>处理A>CK>处理C>处理D;在第17周各处理间的酶活性又略有上升CK的酶活性变成最高,处理A和处理C最低。
2.3.3 4CL酶活性的变化
从图6能看出,不同浓度植物生长调节剂处理对4CL酶活性的影响呈“W”的变化趋势。花后第9周到第17周,CK的4CL酶活性在各处理间始终处于最高值,在第9周,处理A和B的酶活性比处理D和C高;第11周处理B的酶活性最低,处理C和处理D相差不大;第13周4CL酶活性由高到低的顺序分别是:CK、处理A、处理D、处理B、处理C;第15周,CK最高,其他处理的酶活性差距都不大;第17周,所有处理的酶活性都有所回升,处理B、处理C和处理D基本相同,酶活性都较低。
2.3.4 POD酶活性的变化
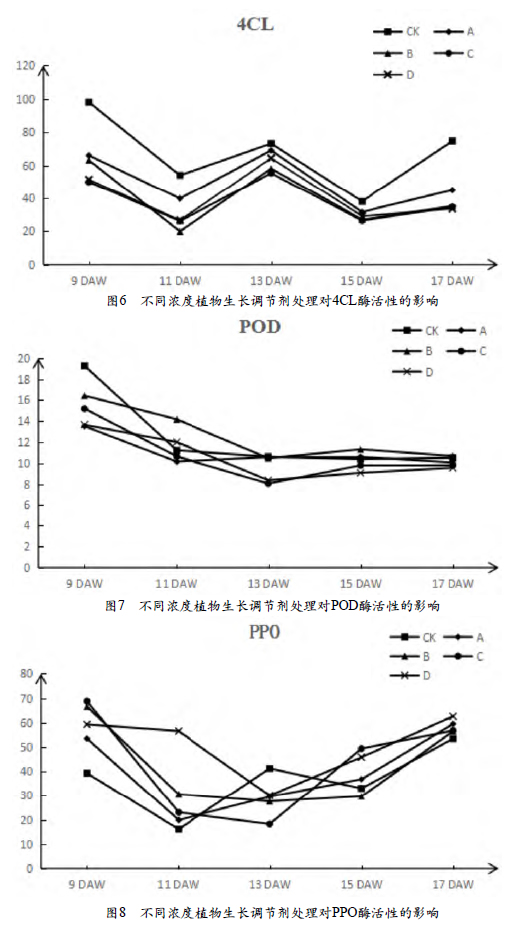
由图7能看出,不同浓度植物生长调节剂处理对POD酶活性的影响呈先下降后持平的趋势。花后第9周,CK的POD酶活性最高,其次是处理B和处理C,处理D和处理A最低;第11周,处理B的酶活性最高,CK、处理C和处理A相差不大,并且活性也较低;第13周,CK>处理A>处理B>处理D>处理C;第15周,处理B的酶活性最大,处理D最小;第17周,各处理POD酶的活性相差不大。
2.3.5 PPO酶活性的变化
由图8能得到,不同浓度植物生长调节剂处理对PPO酶活性的影响整体呈“V型”的趋势。花后第9周,处理C和处理B的POD酶活性最高,其次是处理D和处理A,CK最低;第11周,各处理的酶活性基本都处于下降的趋势,处理D的酶活性基本不变处于最高值,CK处于最低值;第13周,CK的PPO酶活性急剧升高,升到最高,处理D、处理A和处理B相差不大,处于中间位置,处理C酶活性下降到最低;第15周以后,所有处理的酶活性都开始升高,处理C和处理D的酶活性较高,CK、处理A和处理B较低;第17周,所有处理的酶活性相差不大,其中处理D的酶活性最高。
2.4 RNA-seq筛选的与酶活相关基因的qRT-PCR表达
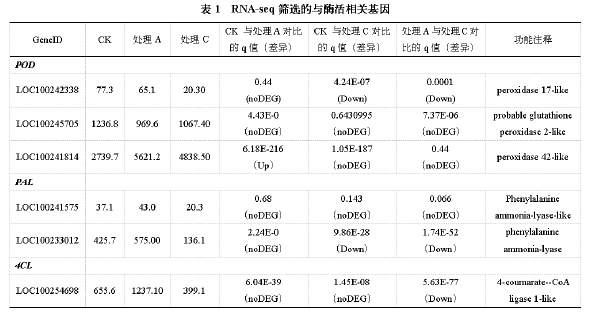
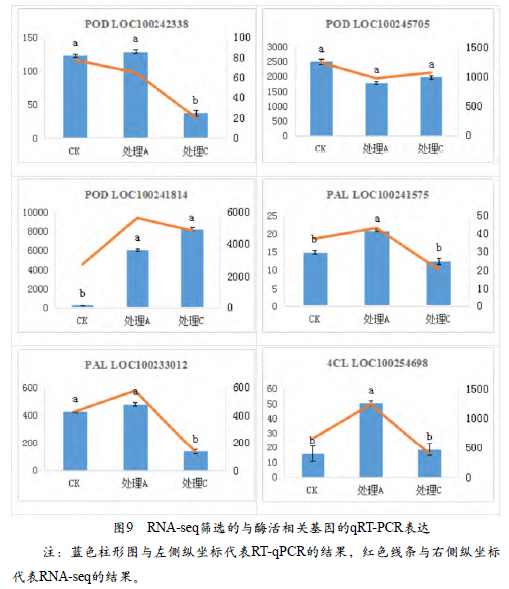
表1是RNA-seq筛选的与酶活相关基因。与POD酶相关的基因有3个,通过不同处理表达量之间的对比,一部分出现上调,一部分出现下调,而LOC100245705没有显著性差异;与PAL酶相关基因有2个,部分处理出现下调;与4CL酶相关基因只有1个。图9是RNAseq筛选的与酶活相关基因的qRTPCR表达与RNA-seq结果的对比,对比的结果基本一致。左一、左二和右一是与POD酶相关基因的表达量,左一是LOC100242338的表达量,CK与处理A表达无明显差异,与RNA-seq相同,处理C与CK和处理A的表达量都有明显差异;右一是LOC100245705的表达量,与RNA-seq结果完全相同,各处理之间均无明显差异;左二是LOC100241814的表达量,处理A和处理C与CK相比基因的表达量都有显著升高。右二和左三是与PAL酶相关基因的表达量,右二图中显示处理A对比CK和处理C的表达量都有显著升高,CK与处理C之间的差异不大;左三是LOC100233012的表达量,与RNA-seq结果基本一致,处理C对比CK和处理A表达量显著下降,CK与处理A的差异不大。右三显示的是4CL酶相关基因LOC100254698的表达量,处理A对比CK和处理C的表达量显著升高,与RNA-seq结果中处理A对比CK有所升高,但差异不明显稍有不同。
3 讨论
葡萄是世界上最受欢迎的餐桌水果之一,葡萄酒也被大量的生产制作。‘阳光玫瑰’葡萄具有很高的可溶性固形物含量和玫瑰型香气,所以在亚洲地区,尤其在中国和日本被大量的种植。在没有经过套袋和植物生长调节剂处理的‘阳光玫瑰’葡萄的表皮上会出现黄褐色的果锈
。套袋能减少果锈在‘翠冠’梨上的发生,但依然存在。GA3和CPPU能增大葡萄、猕猴桃和三倍体枇杷的果实大小;还能提高苹果的单性结实;以及延长葡萄和兔眼蓝莓的货架期。目前,不同浓度的GA3和CPPU处理能减少“阳光玫瑰”葡萄表面果锈的报道还比较少。本文首次通过RNA-seq与木质素和酶活性相结合的方式检测不同浓度的GA3和CPPU处理‘阳光玫瑰’葡萄对果锈的影响。
李国刚等认为果实表面果锈越多,木质素的含量就越高。本研究中就又一次证明了这一观点,CK表面的果锈所占果皮表面积的比例最大,木质素含量也明显高于其他处理;处理C表面的基本没有果锈,木质素含量显著低于其他处理。
木质素是通过相关酶与调控其合成的基因共同作用而进行形成、运输与聚合过程,并且在很多植物中基因家族的成员并不是只有一个。苯丙氨酸解氨酶(PAL)是木质素单体合成过程中是第一个关键酶。在莴苣和非洲菊中PAL酶活性与黄褐色斑点有关,并且与木质素含量呈正相关。但也有研究认为,PAL活性的升高并没有促进木质素的合成。本研究表明在木质素合成关键时期(注:花后14周出现果锈,木质素合成应该在其之前)花后13周,PAL酶活性略微升高,其中CK活性最高,处理C活性最低。与王斌等观点相同,PAL活性与木质素含量呈正相关。4CL活性与PAL相似,在花后13周,各处理4CL活性与木质素含量完全对应,这与Kawaoka等的观点相吻合。据报道,在枇杷果肉和大豆根上发现木质素的合成与POD活性有关,但在本结果中,盛花期11周后,各处理间的POD活性差距很小,基本相同。说明不同浓度的植物生长调节剂对POD活性的影响很小,POD也不是木质素合成主要的限速酶。
本研究中不同处理间C4H活性变化无明显规律,花后第13周,各处理酶活性变化与木质素变化趋势完全不同,这在一定程度上说明C4H可能不是木质素合成代谢的限速酶。上述结果表明在花后第11到13周,PPO活性达到最低,但此时期很可能是木质素合成最多的时期,这与张华云等的PPO活性的降低会导致木质素的合成量减少观点不同。
通过RNA-seq结果,筛选出6个不同处理间与酶活性相关的差异表达基因,经过RT-qPCR验证,这6个基因的RT-qPCR结果与RNA-seq测序结果基本一致,从而证实了基因表达模式的准确性和一致性。在筛选出与POD相关的基因中,LOC100242338和LOC100241814的基因功能是合成过氧化物酶,处理C 显著抑制LOC100242338的表达,这与处理C的木质素含量相对应,各处理的LOC100245705表达量无差异,与POD活性相一致,这说明POD活性可能是受LOC100245705的调控。
LOC100233012的编码苯丙氨酸解氨酶,在LOC100233012的表达量中处理C明显低于其他处理,处理C的PAL活性和木质素含量也是最低,所以LOC100233012很有可能是调控PAL活性的关键基因,从而进一步调控木质素含量,影响果锈的形成。LOC100254698是筛选出于4CL相关的基因,他的功能为4-香豆酰辅酶A连接酶,处理A的LOC100254698表达量显著高于其他处理,这与各处理的4CL活性规律有差异,因此LOC100254698可能不是控制4CL活性的关键基因。
综上所述,处理A能减少果锈的生成,但不能完全清除;处理C可以抑制LOC100233012的表达,进而降低PAL活性,从而抑制木质素的合成来减弱木栓化,最终达到减少果锈的目的;4CL在过程中可能起到辅助作用。因此,在生产上我们可以用25mg·L-1GA3来减轻果锈的发生,同时起到保果的作用,在此基础上再用10mg·L-1CPPU来清除果锈的发生。即通过处理C的方法来抑制LOC100233012的表达,用来减少甚至消除‘阳光玫瑰’葡萄果皮表面的果锈,从而提高其商品价值。