华中西南区栽培岗位
杨国顺 刘昆玉 白描 周敏 郭亮
摘 要:在无公害葡萄和绿色食品葡萄生产过程中,果园土壤重金属元素是产地环境要求检测的一项重要指标。有研究表明,湖南省土壤中存在较为严重的镉污染,土壤镉污染会影响植物的生长、发育,这会对果园的安全生产造成一定的威胁,因此,土壤中的镉元素成为果园生态的主要影响因子之一。镉在葡萄各器官中的分布规律及葡萄对镉胁迫的响应还未见报道。本文以盆栽温克葡萄为材料,进行不同程度Cd2+胁迫处理,研究了Cd在葡萄树体的分布规律和生理特性,结果表明:Cd在葡萄树体的分布规律表现为根>枝条>叶>果实,在高浓度的Cd2+处理下,葡萄根系表现出一定的富集作用,而葡萄果实的Cd含量维持在相对较低的水平。在Cd2+胁迫下,葡萄叶片CAT、SOD、POD等抗氧化酶活性增强,分别在不同浓度达到峰值后开始下降;而葡萄叶片的MDA含量与Cd胁迫浓度呈正相关。低浓度的Cd2+胁迫对葡萄叶片细胞的结构和细胞器无明显损伤症状;当Cd2+浓度为10mg/kg,叶片细胞结构受到破坏,主要表现为膜结构受到破坏、叶绿体变形、类囊体膜膨胀松散以及叶绿体基质解体等现象。
关键词:葡萄;镉; 生理响应; 叶绿体
湖南省被称为“有色金属之乡”,长期的矿产开发活动产生的大量废水、废气和废渣,对周围的水域和土壤环境产生了严重的影响,造成了严重的土壤重金属污染。数据表明,在湖南省长沙市、株洲市、湘潭市等地均存在不同程度的重金属污染,其中以Cd污染较为突出。
土壤Cd污染严重影响着植物的生长、发育,对农作物的生产也造成了严重的威胁。植物受到重金属Cd胁迫,其细胞超微结构通常会受到破坏,这是植物一系列生理活动异常的细胞学基础,会直接反应到植物的代谢、生长等方面。同时,植物体内会诱导产生较多的自由基,自由基的动态平衡遭到破坏。重金属在植物体内积累超过一定的临界值后,细胞膜就会受到伤害,会对植物体内的细胞以及整个植株的水分和离子稳态造成破坏,损伤植物的细胞分子,最终会抑制植物的生长、影响作物的产量和品质。前人对莼菜、小麦、番茄、水稻、烟草等植物的研究表明重金属Cd对植物细胞超微结构的影响主要体现在细胞核、质膜、叶绿体、线粒体、液泡等方面。绿豆、箭舌豌豆、草莓、花生等植物在Cd胁迫下,植物体内抗氧化系统会受到较大的影响,抗氧化酶系统功能紊乱引起的细胞内活性氧自由基动态平衡失调,进而破坏细胞质膜的结构和功能,是重金属对植物产生毒害作用的内在原因。关于镉胁迫下植物生理响应的研究多集中在草本、禾本科类植物,而对果树的研究还较少。湖南省葡萄种植面积已达3.85万hm2,年产量达69.53万t,在全省各个市、州均有种植,而湖南省土壤镉元素已成为果园生态的主要影响因子之一,土壤重金属镉污染可能作为一个潜在因子对葡萄生长、发育构成威胁。
本研究以盆栽葡萄为材料,进行了不同程度的Cd2+胁迫处理,分析了镉在葡萄植株内的分布规律,并对镉胁迫处理下葡萄的生理响应进行了探讨。旨在明确镉在葡萄内的分布特征以及葡萄对镉胁迫的生理耐受性,为葡萄安全生产提供参考。
1 材料和方法
1.1 试验材料
本试验采用1 年生的葡萄扦插苗为材料, 葡萄品种为温克(wink)。盆栽基质由园土、河沙、营养土按3:2:5的比例配置,充分混匀,风干后等量填入盆栽容器中。盆栽基质的理化性状见表1。
1.2 试验设计
葡萄盆栽试验于2014年-2015年在湖南农业大学干杉葡萄基地进行。以黑色塑料桶为盆栽容器,规格为:[30cm(下径)-40cm(上径)] *50cm,容量为21.00kg基质。以浇施土壤的方式进行外源Cd2+处理,外源Cd2+为Cd(NO3)2•4H2O,参照《土壤环境质量标准》(GB15618-1995),设计五个梯度Cd2+处理,分别为0.3、0.6、1.0、3.0、10.0(mg/kg),以清水浇湿土壤为对照(CK),共6个处理,每个处理设置5次重复。
只对盆栽葡萄进行Cd2+胁迫,其它栽培技术均参照常规栽培。取葡萄主梢第5节位叶片为材料进行相关指标测定和观察。
1.3 测定指标及方法
1.3.1 Cd含量的检测
按照GB/T5009.123-2003、GB17141-1997中的规定进行样品预处理和含量测定。采用PE-5000型原子吸收分光光度计检测Cd元素。
1.3.2 抗氧化反应相关指标的检测
氧化反应的相关指标的检测方法参考肖浪涛主编的《植物生理学实验技术》,采用NBT光化还原法测定超氧化物歧化酶(SOD)的活性;采用H2O2一愈创木酚法测定过氧化物酶(P0D)的活性;采用比色法测定过氧化氢酶(CAT)的活性。采用硫代巴比妥酸显色法测定丙二醛(MDA)含量的。
1.3.3 叶绿素含量的检测
叶绿素含量的测定方法参照肖浪涛主编的《植物生理学试验技术》。取新鲜植物叶片,用无菌水洗掉洗净叶面,吸水纸吸干,用9mm打孔器取小圆片材料,打孔时避开叶脉,加入95%的乙醇溶液于暗箱中浸提8-12h,每4h摇动一次,直至小圆片完全变为白色为止,准确定容后即可进行比色、测定分别计算叶绿色a值、b值及叶绿素总量。
1.3.4 叶片细胞超微结构
葡萄叶片用磷酸缓冲液冲洗,快速地切成2mm*2mm的样品块,用2.5%戊二醛进行前固定(6-8h),1M PBS缓冲液(Ph7.2)冲洗3次,15mins/次,3%的锇酸进行固定2.5h,不同浓度的丙酮溶液进行梯度脱水(30%-50%-70%-80%-90%逐级处理,再用纯丙酮作用3次,30mins/次),Epon812进行梯度渗透、包埋,再进行修块、定位、切片、铜网捞片等,用柠檬酸铅、醋酸铀双重染色后,于TEMH-7000型透射电镜下观察、拍照,透射电镜观察超薄切片时,条件为加速80kv,束流50LA,聚光镜孔径200Lm,物镜孔径40Lm(Jin et al.,2008)。
2 结果与分析
2.1 Cd 在温克葡萄树体内的分布特征
在低浓度的Cd胁迫下,温克葡萄根系、枝条和叶片的中的Cd含量差异不显著;当Cd2+浓度为3.0mg/kg时,根系和枝条中Cd含量显著提高;Cd2+浓度为10.0mg/kg时,根系和枝条中的Cd含量分别达到8.170mg/kg、0.290mg/kg;果实中Cd含量与Cd胁迫浓度无相关关系,最低值为0.010mg/kg,最高值为0.021mg/kg。温克葡萄根系对Cd的吸收能力较强,在高浓度Cd处理下,根系中的Cd浓度为8.170mg/kg,根系表现出对Cd的富集作用,但温克葡萄果实Cd含量没有明显的增加,始终保持在较低的水平,Cd在温克葡萄各器官中的分布规律大致表现为:根>枝条>叶片>果实。
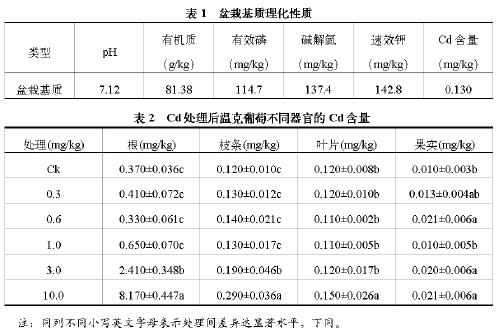
2.2 Cd胁迫对葡萄生理特性的影响
2.2.1 Cd胁迫对温克葡萄叶片抗氧化反应的影响
温克葡萄在Cd胁迫下,叶片POD活性增强,浓度为1.0mg/kg时,达到最大值,为对照的1.75倍,随着浓度的增加有所下降;叶片的CAT活性在Cd2+浓度为3.0mg/kg时出现最大值,为对照的5.92倍,当浓度继续升高,叶片的CAT活性开始下降;Cd胁迫下,叶片的SOD活性均高于对照,Cd2+浓度为0.6mg/kg时,叶片的SOD活性是对照的2.63倍;叶片的MDA含量随着处理浓度提高不断上升,MDA含量最高值为9.19umol/g,显著高于对照,是对照的2.13倍。结果表明,Cd胁迫条件下,温克叶片的POD、CAT、SOD活性均高于对照,且在一定浓度范围内,与Cd处理浓度呈正相关,随着处理浓度继续提高,活性开始下降;叶片的MDA活性与Cd处理浓度成正相关。
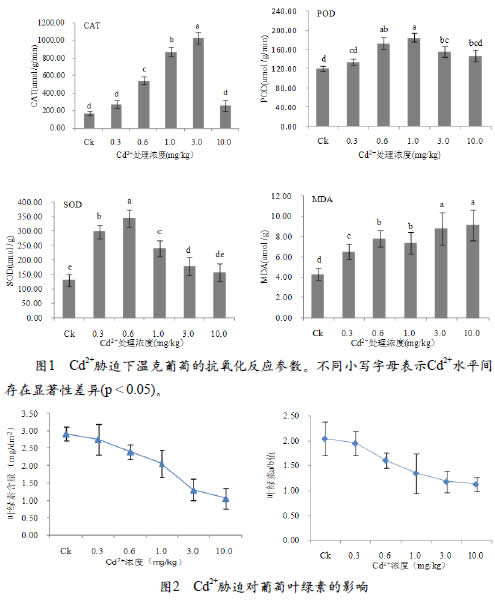
2.2.2 Cd胁迫对葡萄叶片叶绿素的影响
在C d 处理下, 葡萄叶片的叶绿素含量受到显著影响,温克葡萄叶片叶绿素含量为1.07±0.29~2.92±0.19mg/dm2,温克葡萄叶片的叶绿素含量随着Cd2+浓度的增加而降低,10mg/kg的Cd2+处理下,温克葡萄叶片叶绿素含量降低了63.4%;温克葡萄叶片叶绿素a/b值为2.04,在Cd胁迫下,温克葡萄叶片a/b值降低,Cd2+浓度为10mg/kg时,叶绿素a/b值最低为1.13。结果表明,葡萄叶片的叶绿素含量及叶绿素a/b值均与Cd2+浓度呈负相关。
2.2.3 Cd胁迫对葡萄叶片细胞结构的影响
本研究分别对温克葡萄叶片细胞结构进行了透射电镜观察,用于制作切片的葡萄叶片采自Cd2+处理浓度为0mg/kg(ck)、0.3mg/kg(低)、1.0mg/kg(中)、10.0mg/kg(高)等四个处理的植株,将对照和高、中、低浓度的Cd胁迫叶片进行对比观察,温克葡萄叶片细胞结构见图3。
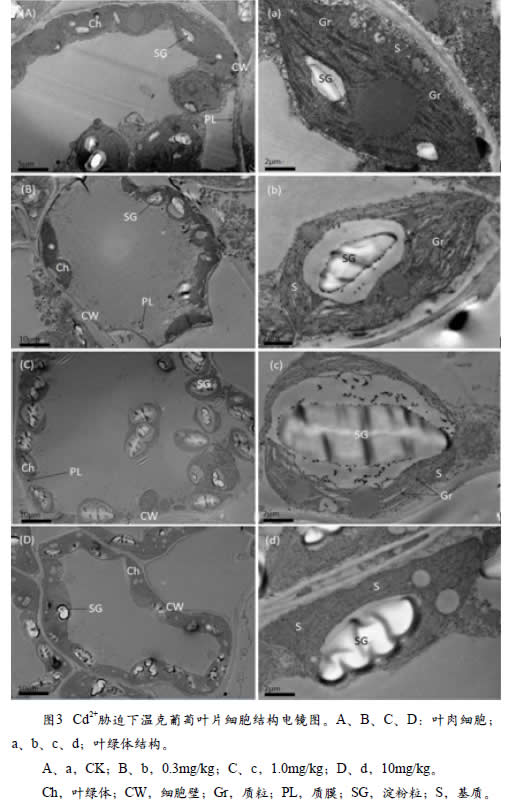
温克葡萄的对照叶片细胞中,细胞壁、细胞膜结构完整,叶绿体呈现纺锤形,分布于细胞边缘,基粒片层排列有序,基粒片层和类囊体膜系统清晰,叶绿体内部包含淀粉粒。低浓度Cd处理下,细胞形态未见明显变化,随着Cd浓度的增加,细胞膜结构被破坏,细胞膜变薄,叶绿体内部结构受到不同程度的破坏,细胞叶绿体变圆、变小,而且淀粉粒数量增多;Cd浓度为10.0mg/kg时,类囊体膜结构松散,基粒排列无序,叶绿体内部内部出现空泡化,叶绿体膜受损,叶绿体发生解体,叶绿体基质连成一体。
3 讨论
3.1 Cd在葡萄植株内的分布特征
植物对重金属Cd的吸收与植物品种、土壤中的Cd含量以及土壤环境密切相关(赵中秋等, 2005)。Cd作为一种植物生长的非必需元素,其进入植物体内,可能随着水分吸收或钙离子通道等途径被动地进入植物体内,进入植物后主要滞留在根部的质外体中,如导管壁、木质部细胞壁中,并被细胞壁中的果胶酶、纤维素吸附,其中还有少量Cd会积累在液泡中,这样就限制了大部分Cd的移动性,少量的Cd可能在蒸腾作用下,随着水分流动,通过导管向地上部分运输,依次经过茎杆、枝条后,被这些部位的纤维素吸附,到达果实的Cd非常少了,表明植物的根系和茎杆对Cd有天然的过滤和固定作用,能够有效的阻止Cd向植物地上部分迁移,减轻Cd的生物毒性。
葡萄根、茎、叶、果实等器官中Cd含量可以反映葡萄对Cd的吸收能力和Cd在植物体内的分布特征。Cd在葡萄树体的分布规律表现为根>枝条>叶>果实,表明葡萄树体吸收的Cd大部分积累在葡萄的根系,随着浓度的增加,才有少量的Cd会往上运输到枝条和叶片,最后才到葡萄果实,即使在高浓度的Cd2+处理下,葡萄果实的Cd含量也维持在相对较低的水平。这与苹果、阳桃、黄皮、龙眼、玉米、大豆等作物对Cd的积累特征相类似。
3.2 Cd胁迫对葡萄生理特性的影响
Cd对植物产生的毒害作用,与植物体内的自由基的变化有密切关系。当植物受到Cd胁迫时,会诱导产生较多的自由基,植物体内的抗氧化系统会自发地清除过多的自由基,这个系统包括酶(SOD、CAT、POD等)和非酶两个系统。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶( CAT)是植物抗氧化酶系统的主要成员,它们在植物适应多种逆境胁迫中发挥着重要作用。在逆境条件下,为了减轻毒害作用,植物机体会产生较高的抗氧化酶活性来清除体内多余的过氧化自由基。在植物体内,SOD能够有效地是清除O2˙-,可将其歧化H202,后者的氧化作用较弱;而CAT和POD这两种酶则可以进一步将H202转化为H2O和O2,从而减少O2˙-和H202的积累。如果植物体内的O2˙-和H202含量过高,不能被及时清除,这二者就可能通过Fenton反应而生成OH, 其生物毒性更强,进而对蛋白质、膜脂及核酸产生伤害,已有研究表明,植物体内的抗氧化酶活性与O2˙-和H202含量有显著的正相关,有学者建议植物的抗氧酶活性的变化水平可以反应植物对污染胁迫耐受指标。MDA是反应植物细胞膜脂过氧化作用的一个重要指标,膜脂过氧化作用是发生在细胞膜上的一系列活性氧反应,是植物受重金属胁迫毒害的内在机理。Cd2+胁迫会导致植物的自由基过氧化作用带来损伤,这样会使植物体内膜脂过氧化作用加强,植物体内的自由基活动的状态一定程度上可通过MDA含量来反映,MDA含量越高,植物所受胁迫产生的自由基含量越高,表明植物所受毒害作用越严重。番茄受到Cd2+胁迫时,其叶片细胞超微结构受到破坏,表现为叶绿体变形、扭曲。水稻在Cd2+处理下,对叶片细胞产生了影响,其叶绿体片层空泡化,叶绿体基质变淡。Rodrigo等用Cd2+处理Gracilaria domingensis 后,发现叶片细胞中叶绿体的类囊体遭受到损伤,同时出现了质粒小球,随着Cd2+浓度的增加,叶绿体受到的伤害越严重。
本试验中,盆栽葡萄在Cd胁迫条件下,叶片的POD、CAT、SOD等活性有所提高,在一定的Cd2+胁迫范围内,葡萄叶片的这三种抗氧化酶活性与Cd2+浓度呈正相关,随着浓度的继续提高,抗氧化酶活性会出现下降,可能是因为高浓度的Cd2+胁迫对叶片细胞造成了不可逆转的损伤,这与油菜、萝卜等植物在重金属胁迫下的生理特性相似。葡萄叶片的MDA含量与Cd2+胁迫浓度呈正相关,表明Cd2+胁迫会导致膜脂过氧化作用,这与绿豆、箭舌豌豆等植物在重金属胁迫下的生理特性相似。Cd2+处理对温克葡萄的茎叶等外观形态上没有造成明显的伤害,不同Cd2+浓度处理后,葡萄的生长发育状况都比较一致,但对葡萄叶片的细胞结构造成了一定的破坏,其中最为明显的是细胞膜结构和叶绿体的变化。低浓度的Cd2+胁迫对葡萄叶片细胞的结构和细胞器无明显损伤症状,当Cd2+浓度为10mg/kg,叶片细胞的细胞膜结构受到破坏,主要表现为膜结构受到破坏、叶绿素变形、类囊体膜膨胀松散以及叶绿体基质解体等现象。
叶绿体是对Cd2+胁迫最敏感的细胞器之一,可能是由于细胞渗透压的变化引起叶绿体膜系统遭到破坏,导致叶绿体解体,而叶绿体中的淀粉粒的变化可能是在光合作用合成的糖类未能正常输出而形成的淀粉累积过多而成。叶绿体是植物进行光合作用的场所,是植物的能量工厂,叶绿体对重金属胁迫比较敏感。当叶绿体基质类囊体膜和叶绿体被膜受到破坏时,其光合功能将受到影响,甚至失去光合功能。细胞膜是细胞内外物质交换的门户,对外来物质具有选择透过性,既能选择性地透过细胞代谢所需的矿质元素,又能阻止一些有害物质的进入,维持着细胞内相对稳定的环境。受到重金属胁迫,往往会发生膜脂过氧化作用,使细胞膜结构或功能先受到破坏,其产物之一就是MDA,因此,MDA含量可以用来反映细胞膜脂过氧化作用的程度,本研究的结果佐证了这一点,重金属对茶树、水稻、白菜等植物的细胞也造成了类似的伤害。葡萄在低浓度的Cd2+胁迫下,叶片抗氧化酶活性提高,能够通过自身的抗氧化系统来清除体内的自由基;而随着胁迫浓度增加,葡萄的抗氧化系统功能紊乱,对自由基的清除能力下降,细胞内过多的自由基就会加剧膜脂过氧化作用,对细胞质膜产生毒害作用。