华东华南栽培岗位
辛守鹏 刘帅
陶建敏
光合作用是植物生长发育的基础和生产力的决定性因素,也是衡量外界对植物影响的重要指标,较高的光合能力是葡萄果实高产的前提。植物净光合速率光响应曲线描述的是光量子通量密度与植物净光合速率之间的关系,
即净光合速率随光照度的变化特征,因而光响应曲线则成为衡量叶片光合能力的重要指标。为准确细致研究叶片光响应曲线就需要对其进行数学化拟合,但在很多的研究论文表明,不同拟合模型对植物叶片光响应曲线拟合的适用性是不同的。植物叶片的多样性决定了叶片光响应曲线拟合模型的差异性,不同的模型对同一物种植物光响应曲线拟合的适用性会有差异,同一种模型对于同一物种的不同品种甚至同一品种植物不同部位叶片的适用性也极有可能不相同。因此,在对植物叶片光响应曲线进行拟合时,需要充分考虑到所使用的拟合模型对于所研究叶片光响应曲线拟合的适用性,选择适合的光响应曲线拟合模型是准确衡量叶片光合能力的先决条件。所以,筛选出植物最适合的光响应曲线拟合方法对准确研究植物的光合特性有重大意义。利用二项式回归、直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数函数模型5种拟合方法分别对正常情况下和用CPPU处理后阳光玫瑰葡萄果实邻近叶光相应曲线拟合,比较5种方法拟合后的光响应曲线及相关参数的差异,筛选出最适合的光响应曲线拟合方法,为后续光合特性的研究提供理论基础。
1 材料与方法
1.1 试验设计
试验于2014年在南京农业大学汤山葡萄试验基地进行,平棚架“H”型整形避雨栽培,南北走向,行株距为6.0m×3.0m。试验品种为6年生阳光玫瑰葡萄,以长势相近的植株为试材,常规方法进行田间土肥水管理及病虫害防治。处理前对花序进行修剪,除去穗肩上2到3个分枝,留花序长约4 cm,约花后一周(5月20日)处理。处理时将花序在清水(编号CK)、 20mg/LCPPU(编号AK)药液中浸泡2~3s,每个处理3棵树, 共两个处理。 6月20日左右,选择晴天的9:00~11:30,从每个处理中中选取长势一致无其他叶遮挡的果实邻近叶片(果实上部1~2片叶),用美国LI-COR公司生产的LI-6400便携式光合测定仪进行测定,重复3次,取平均值。外界通路条件下流速设定为500μmol•s-1,在0~2000μmol•m-2•s-1光照范围内设置12个光照强度(0、20、50、100、200、400、600、800、1000、1200、1600、2000μmol•m-2•s-1),测定各处理叶片的净光合速率(Pn)的光响应曲线。
1.2 光响应曲线的拟合方法
(1)二项式回归:二项式回归法是以净光合速率(Pn)和光合有效辐射(PAR)的成对值进行二元回归,Pn最大时即为最大净光合速率 (Pnmax,μmol•m-2•s-1),此时的PAR为光饱和点 (LSP,μmol•m-2•s-1),PAR为0时的Pn为暗呼吸 (Rd,μmol•m-2•s-1)。
(2)直角双曲线模型表达式(图a):式中:I为光合有效辐射;Pn(I)为光强为I时的净光合速率;α为表观量子效率;Rd 为暗呼吸速率。
(3)非直角双曲线表达式(图b):式中:k为非直角双曲线的曲角,其他参数意义同上。
(4)直角双曲线修正模型表达式(图c):式中:β为修正系数,γ是一个与光强无关的系数,其他参数意义同上。
(5)指数函数模型表达式(图d):式中:e为自然数对数底,其他参数意义同上。
1.3 数据处理
采用二项式回归、直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数函数模型对叶片光响应曲线进行非线性拟合。用SPSS16.0统计软件和Excel2007软件对数据进行统计分析。
2 结果与分析
2.1 二项回归对光相应曲线的拟合
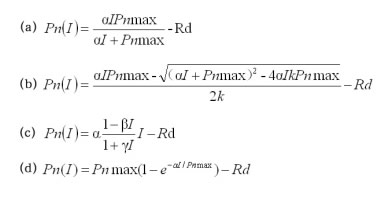
从用二项式回归拟合的AK和C K的光响应曲线可以看出(图 1),AK与CK的净光合速率(Pn)实测值的变化趋势都是随着光合有效辐射(PAR)的增强而增大,当光合有效辐射达到一定强度后,净光合速率Pn增加缓慢,基本没有变化,更没有降低;而由二项式拟合的AK和CK的光响应曲线得到的光补偿点(LCP)为负值,暗呼吸速率为正值,这与实际情况明显相悖,与拟合多种植物的结果类似,同时该曲线在PAR达到一定强度值后有明显下降的趋势,这也与实测值明显不符;但二项式回归方程是一个有极值的函数,通过其可以得到光饱和点(LSP)及最大净光合速率(Pnmax)。二项式回归拟合的AK和CK的光响应曲线虽能得到相关参数,但得到的部分参数数值与实测值差异较大。
2.2 直角双曲线模型对光相应曲线的拟合
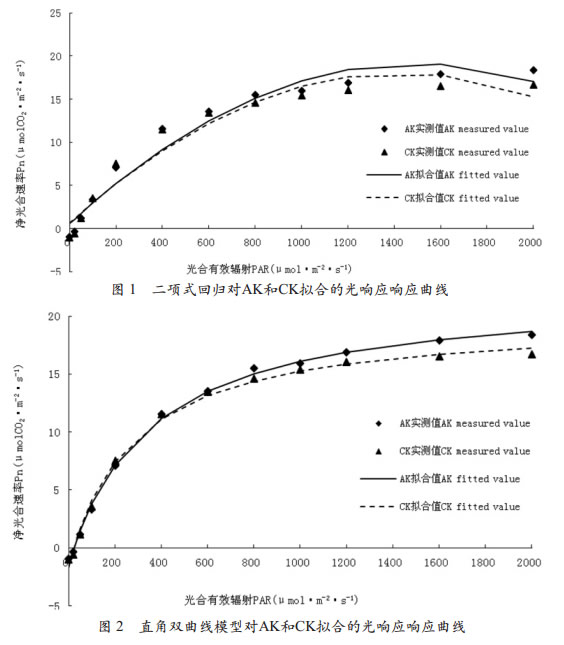
从用直角双曲线模型拟合的AK和CK的光响应曲线可以看出(图 2),利用直角双曲线模型拟合光响应曲线效果较好,但在A K处理光合有效辐射为800μmol•m-2•s-1处,拟合值明显低于实测值,同时在光合有效辐射达到2000μmol•m-2•s-1时,拟合值又高于实测值;在CK处理1600~2000μmol•m-2•s-1范围内,拟合值高于实测值,同时直角双曲线模型拟合的AK和CK的光响应曲线不能直接给出光饱和点。
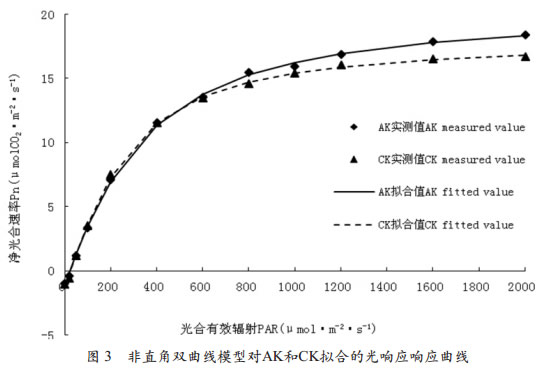
2.3 非直角双曲线模型对光相应曲线的拟合从用非直角双曲线模型拟合的AK和CK的光响应曲线可以看出(图 3),利用非直角双曲线模型拟合的AK和CK的光响应曲线拟合效果较好,与实测值极为接近,特别是对低光合有效辐射区和高光合有效辐射区范围内的实测值有很好的拟合效果;但非直角双曲线模型拟合的AK和CK的光响应曲线与直角双曲线模型拟合的AK和CK的光响应曲线存在同样的缺陷,无法由曲线直接得到光饱和点。
2.4 直角双曲线修正模型对光响
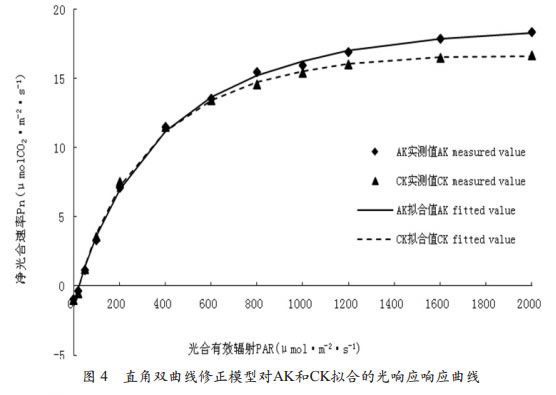
应曲线的拟合从用直角双曲线修正模型拟合的AK和CK的光响应曲线可以看出(图 4),利用直角双曲线修正模型对AK和CK拟合的光响应响应曲线拟合效果总体也很好,与实测值变化趋势基本相符,通过曲线可以得到光饱和点;但同时我们也能看出,利用非直角双曲线模型对AK和CK拟合的光响应曲线呈现出先上升后下降的规律,当光合有效辐射在1600μmol•m-2•s-1以下时,随着光合有效辐射的增加,拟合曲线呈上升趋势,而在光合有效辐射达到1600μmol•m-2•s-1以上时CK光响应曲线呈下降趋势,这与实测值随光合有效辐射增强不断增大的事实不相一致。
2.5 指数函数模型对光相应曲线的拟合
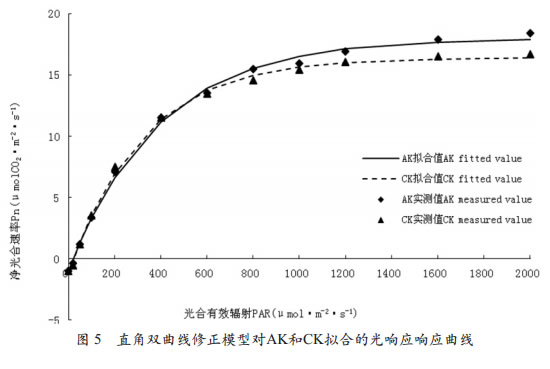
从用指数函数模型拟合的AK和CK的光响应曲线可以看出(图 5),利用指数函数模型对AK和CK拟合的光响应响应曲线拟合效果较好,拟合值与实测值变化趋势基本相符,但通过曲线不能得到光饱和点;同时我们可以看到,当光合有效辐射400 mol•m-2•s-1以下时拟合值明显低于实测值,而当光和有效辐射在600~1200 mol•m-2•s-1之间时,拟合值又高于实测值,再当1600~2000mol•m-2•s-1的范围内时,拟合值又低于实测值,总体而言,指数函数模型拟合的光响应曲线拟合值与实测值有较大的差异。
2.6 5种光响应曲线拟合方法相关参数的比较
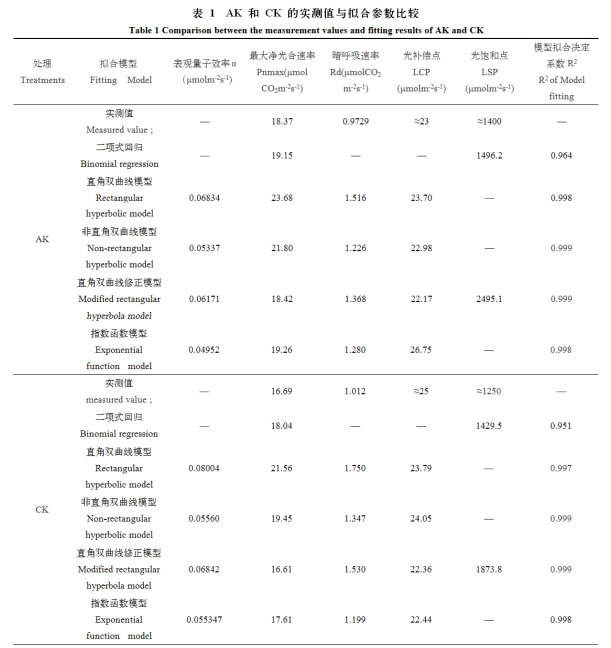
通过对5种拟合方法拟合的AK 和 CK光相应曲线相关参数与实测值对比分析可以看出(表 1),二项式回归拟合方法得到的最大净光合速率与光饱和点与实测值较为接近,但做得到的暗呼吸速率和光补偿点与事实相悖,没有实际意义;直角双曲线模型和非直角双曲线模型拟合得到的最大净光合速率、暗呼吸速率和光补偿点与实测值都比较接近,而非直角双曲线模型拟合得到数值比直角双曲线更接近实测值,且非直角双曲线拟合决定系数比直角双曲线高,拟合效果更好,但直角双曲线模型和非直角双曲线模型都是没有极值的函数,因此无法求取植物饱和光强;而直角双曲线修正型拟合得到最大净光合速率和光饱和点与实测值几乎一致,但暗呼吸速率和光饱和点明显高于实测值,差异较大,同时拟合曲线随着光合有效辐射的增强先增加后降低的趋势,与实测值随光和有效辐射增强不断增大的事实不相符;由指数函数模型拟合的光响应曲线得出的最大净光合速率、暗呼吸速率与实测值较为接近,但AK拟合光响应曲线的光补偿点明显高于实测值,而AK拟合的光响应曲线光补偿点又低于实测值,且模型拟合决定系数小于由非直角双曲线模型和直角双曲线修正模型拟合决定系数。
众多有关表观量子效率的研究表明,植物表观量子效率的理论值在0.08~0.125之间,但在实际测量条件下,实测值会小于理论值,其范围在0.04~0.07。由表1 可以看出,各个模型拟合光响应曲线得到的表观量子效率均处于实测值范围以内,同时模型拟合系数都高于0.95,说明5种光响应曲线拟合方法对AK与CK的拟合效果较好。但模型拟合系数的高低不是唯一决定因素,光合速率、表观量子效率、光饱和点、光补偿点等可由植物光响应曲线估算的光合参数是植物重要的生态生理参数,最终需要综合光响应曲线相关参数与实测值差异大小,选择出最适合的光相应曲线拟合模型。
3 结论与讨论
试验中的实测值是光合有效辐射在0~2000 mol•m-2•s-1范围内的几个点,也可以说测出来的也仅仅是光合有效辐射在0~2000
mol•m-2•s-1范围内时整个光响应曲线的一段,光合有效辐射超过2000 mol•m-2•s-1的净光合速率没有测定,这是因为自然状态下的光合有效辐射很难达到2000
mol•m-2•s-1以上,所以能对光合有效辐射在0~2000 mol•m-2•s-1范围内光响应曲线进行最精确数学化拟合的才是最适合的拟合方法,更具有实际应用意义。通过对实测值的观察,AK和CK的净光合速率随着光合有效辐射的增强而增加,没有下降,与李雅善等在对赤霞珠葡萄进行光相应曲线拟合时赤霞珠在高光强下并没有出现光抑制的现象相一致,与惠竹梅等高光强下葡萄叶片出现光抑制现象的研究结果有所不同。对于像阳光玫瑰、赤霞珠等葡萄叶片在2000 mol•m-2•s-1时没有出现光抑制现象,李雅善等得出最适合模型是有极值的直角双曲线修正型,本人认为不妥。直角双曲线修正模型方程本身决定其有极值,在叶片光合能力较大时(如本试验AK),用直角双曲线修正型的出的光饱和点大于2000 mol•m-2•s-1,与李雅善在赤霞珠上得出的结果类似,但当叶片光合能力较弱时(如本试验CK),用直角双曲线修正型拟合得出的光饱和点小于2000 mol•m-2•s-1,明显与事实不符,而非直角双曲线能很好的模拟植物不出现净光合速率随光强增加而降低的光响应曲线。
对于非直角双曲线模型拟合参数光饱和点可以用如下方式获得,先假定光合有效辐射在0~200µmol•m-2•s-1时Pn (I)观察值为一条直线,该直线与Y=Pnmax直线相交, 交点所对应于X轴的数值即为光饱和点。但本人用这种方法得出出的光饱和点明显小于实测值。在较弱光强下(PAR≤200μmol•m-2•s-1),净光合速率对光合有效辐射的响应情况即表观量子效率,反映出叶片对光合有效辐射的利用效率。通常对PAR≤200μmol•m-2•s-1的净光合速率实测值进行线性拟合,由直线斜率得出表观量子效率。而在本文中,本人认为非直角双曲线模型拟合优度极高,所有就由方程直接得出暗呼吸速率、光补偿点等相关参数,而没有对PAR≤200μmol•m-2•s-1的净光合速率进行线性拟合获得,所以得出的表观量子效率、暗呼吸速率、光补偿点等参数准确精度尚待进一步进行验证。
光响应曲线反映了在一定光合有效辐射强度下植物潜在的光合能力,因而光响应曲线的数学花拟合也是研究光响应曲线不可缺少的一环。本文利用5种常用植物光响应曲线拟合方法对AK和CK果实邻近叶光相应曲线分别拟合,得到了用5种光相应曲线拟合方法拟合成的5组光相应曲线,5种光响应曲线拟合方法的拟合结果存在较大差异。
非直角双曲线模型的拟合方程无极值,拟合曲线为一条渐近线,与实测值没有出现光抑制现象的事实相吻合。因此,非直角双曲线模型是最适合阳光玫瑰葡萄果实邻近叶光响应曲线的拟合方法。