加工与综合利用
兰义宾
1 研究目的与意义
单宁作为葡萄酒的“骨架”,其含量不仅决定着葡萄酒的涩感强弱,也影响了葡萄酒的颜色,对葡萄酒的品质至关重要,因此在酿造过程中被广泛添加和使用。这些商业缩合单宁依其来源可分为葡萄皮单宁和葡萄籽单宁,两类单宁结构不同,因而使用后对葡萄酒颜色和口感的影响各有优劣。明确不同类型葡萄单宁添加对葡萄酒感官属性的影响,并且针对不同陈酿条件和不同基质葡萄酒使用最合适的葡萄单宁,最大限度改善葡萄酒的颜色和口感,是酿酒师与葡萄酒企业面临的关键生产问题。‘赤霞珠’作为我国第一大酿酒葡萄品种,在诸多葡萄酒产区得以广泛种植,用以生产和酿造优质葡萄酒。‘马瑟兰’是‘赤霞珠’和‘歌海娜’的杂交品种,于2001年引入中国,因其品质优良,抗病性强的特点,种植面积逐年增加。近年来,外源葡萄单宁已经成为在葡萄酒酿造阶段广泛使用的加工助剂,其对于葡萄酒品质的影响也成为研究热点之一,尽管越来越多的研究和生产现象发现外源葡萄单宁会对葡萄酒的颜色和口感产生影响,但不同来源的葡萄单宁对葡萄酒品质的贡献尚未明晰。
基于此,本研究分别于2018年和2019年向橡木桶陈酿前的‘赤霞珠’和‘马瑟兰’葡萄酒中添加不同来源的葡萄单宁(葡萄皮单宁、葡萄籽单宁和葡萄皮籽混合单宁),利用高效液相色谱-三重串联四极杆质谱(HPLC-QqQ)分析葡萄酒酚类物质轮廓,利用高效液相色谱-荧光检测(HPLC-FLD)解析葡萄酒中缩合单宁含量,结合CIELab参数和ASPI表征不同酒样的颜色与涩感特征差异,旨在探究添加不同来源葡萄单宁对橡木桶陈酿葡萄酒颜色和涩感的影响,以期为橡木桶陈酿过程外源添加单宁调控葡萄酒品质提供理论依据和生产实践指导。
2 材料与方法
2.1 葡萄酒样品
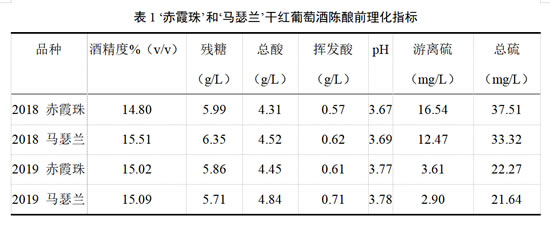
葡萄酒样品为新疆玛纳斯产区中信国安葡萄酒业有限公司于2018年和2019年采摘并酿造的‘赤霞珠’(Cabernet Sauvignon)和‘马瑟兰’(Marselan)干红葡萄酒,所有干红葡萄酒均以成熟度良好的葡萄以工业化葡萄酒的酿造方式进行酒精发酵和苹果酸-乳酸发酵。2018年和2019年‘赤霞珠’和‘马瑟兰’葡萄酒在发酵结束后采用中度烘烤的法国圣哥安橡木桶进行陈酿。根据供应商所建议的添加量,每个品种的葡萄酒均设置在陈酿前添加200 mg/L葡萄籽单宁(ST)、200 mg/L葡萄皮单宁(SKT)、200 mg/L葡萄皮籽混合单宁(MT,混合比例1:1)的3种处理组和不添加单宁的对照组,陈酿周期为12个月,分别在陈酿初始点第0个月(0M)、陈酿6个月(6M)及陈酿12个月(12M)取样进行分析。所有处理组和对照组均设置2个重复,橡木桶陈酿温度控制在14-16℃,湿度控制在65-75%。入桶前的葡萄酒基本理化指标见表1。
2.2 葡萄酒中酚类物质的测定
采用高效液相色谱-三重串联四级杆质谱联用(HPLC-QqQ-MS/MS)检测葡萄酒中的单体花色苷、花色苷衍生物与非花色苷酚。葡萄酒上样前用0.22 μm聚醚砜滤膜过滤。使用Agilent 1200-6410B、Poroshell120EC-C18色谱柱(150 mm×2.1 mm×2.7 µm, Agilent Technologies, US)和电喷雾离子源(ESI)完成检测。喷雾电压为±4000 V。雾化气为高纯氮气,其雾化压力为35 psi。干燥气为高纯氮气,其温度为350℃,流速为12 L/min。扫描模式为多反应监测(MRM)。
利用Masshunter软件(QQQ定量分析)积分峰面积,并将其作为定量依据。通过不同梯度的标准品物质峰面积与浓度的线性拟合曲线进行定量,其中花色苷及其衍生物由二甲花翠素-3-O-葡萄糖苷的标准曲线进行半定量。
2.3 葡萄酒中缩合单宁的测定
采用SPE结合Agilent 1100系列HPLC系统配备G7121A荧光检测器仪分析缩合单宁的浓度及聚合度。将5 mL酒样加至Oasis HLB固相小柱。使用20 mL混合溶液(丙酮/水/甲酸, 80:19.5:0.5, v/v/v)洗脱缩合单宁,收集洗脱液并于高通量旋转蒸发仪中除去丙酮,随后对剩余洗脱液进行冷冻干燥。完毕后将其重新溶解于1 mL模拟酒溶液中,进行后续分析。采用Develosil Diol 100 Å(250 mm×4.6 mm, 5 µm)色谱柱分离不同聚合度的单宁。流动相A为含2%(v/v)乙酸的乙腈溶液,流动相B为甲醇/水/乙酸溶液(95:3:2, v/v/v),洗脱程序如下:B相在3 min内升至7%,在60 min内升至37.6%,在63 min内升至100%,63 min至70 min保持100%,70 min至76 min降至7%,后运行时间为10 min。流速为1 mL/min, 柱温保持在35℃。荧光检测器的激发波长和发射波长分别为230 nm和321 nm。负离子模式采集质谱图,干燥气温度300℃,流速为8 L/h。雾化器压力为35 psi。碎裂电压65 V,离子扫描范围为100 m/z至3200 m/z,扫描时间2 s。使用表儿茶素标准品溶于模拟酒溶液配制标准曲线对缩合单宁进行定量。
2.4 葡萄酒颜色参数的测定
使用CM-3700d分光测色计测定葡萄酒的颜色参数(亮度L*、红/绿色调a*、黄/蓝色调b*)。每个样品进行三个独立技术重复。葡萄酒样品经0.22 μm聚醚砜滤膜过滤后,选择光径为2 mm的玻璃比色皿,利用分光测色计进行检测。分光测色计选择D65标准光源照明、10°的视野条件及孔径为8 mm的MAV目标罩,每次测量前需行白板校准和零位校准。颜色参数由SpetraMagicNX获得,其色板图利用Colortell在线网址(https://www.colortell.com/)绘制。
2.5 葡萄酒涩感强度的测定
利用人工唾液指数沉淀法评价各处理组酒样的涩感强度。人工唾液缓冲溶液含有19.3 mg/L溶菌酶、103.2 mg/L牛血清蛋白、145.7 mg/L乳铁蛋白、233.6 mg/L聚L-脯氨酸、425.8 mg/L α-淀粉酶、0.441 g/L二水合氯化钙、0.477 g/L 氯化钾、0.877 g/L氯化钠、1.369 g/L三水合磷酸氢二钾和5.208 g/L碳酸氢钠,pH为6.6。将人工唾液与酒样按照8:1的体积比混合,混合物在37℃水浴孵育5 min后在4℃,10,000 g下离心2.5 min,取上清液用于高效液相色谱(HPLC)和聚丙烯酰胺凝胶电泳(SDS-PAGE)分析。
采用HPLC法测定聚-L-脯氨酸的损失率。使用Agilent 1100系列的HPLC系统与Zorbax 300SB-C18色谱柱(4.6×250 mm, 5 µm, Agilent Technologies, US)进行检测。进样量为80 μL,色谱柱柱温为40℃。流动相A为含有0.2%的三氟乙酸水溶液,流动相B为含有0.2%三氟乙酸的70/30(v/v)甲醇/乙腈溶液。梯度洗脱程序为:10%~22%B,0~10 min;22% B,10~15 min;22%~28%B,15~23 min;28%B,23~25min;28%~60%B,25~40 min;60%~100%B,40~40.1 min;100%B,40.1~50 min。流速为0.4 mL/min。紫外检测信号设置为214 nm。利用Agilent 1100工作站(Data Analysis)积分峰面积作为定量依据。配制不同梯度(0.2、0.5、1.0、2.0、2.5、4.0和5.0 g/L)的单宁溶液用于校准曲线。
采用SDS-PAGE法测定α-淀粉酶、乳铁蛋白、溶菌酶和牛血清白蛋白的损失率。SDS-PAGE分析在Mini-Protean Tetra Vertical Electrophoresis Cell仪器上进行。上样量为30 μL,电压设置为140 V。染色液为考马西亮蓝(R-250),染色时间为30 min;脱色液为10%的醋酸溶液,脱色时间为12 h。使用GelDoc 2000凝胶成像系统记录,记录蛋白质电泳的图像。通过ImageJ 1.53a软件分析蛋白的灰度损失。
人工唾液沉淀指数以各蛋白损失率计算,公式如下:
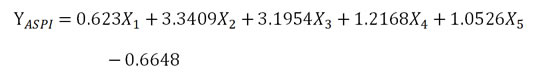
3 结果与讨论
3.1 不同来源葡萄单宁对陈酿过程中‘赤霞珠’葡萄酒颜色和涩感的影响
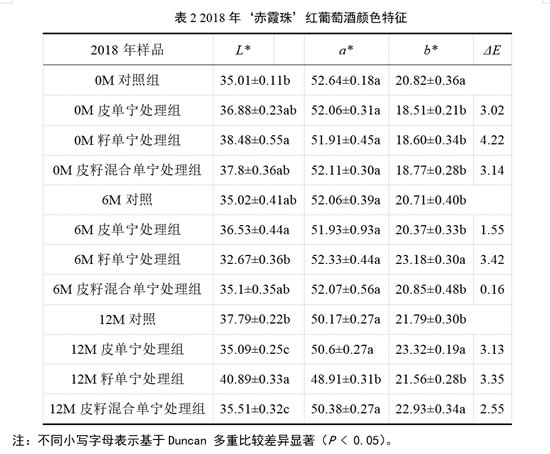
对于2018年‘赤霞珠’干红葡萄酒而言,由表2可知,在经历了12个月桶储陈酿后,籽单宁处理组的酒样L*值最高,且显著高于其他处理组,表明其酒样的颜色最浅。对干红葡萄酒的a*值而言,仅在陈酿12个月时,籽单宁处理组的葡萄酒a*值显著低于其他处理组。而对于酒样的b*值,在桶储陈酿6个月后,籽单宁处理组的b*值显著高于其他处理组,当陈酿12个月时,皮单宁处理组b*值最高,且显著高于对照组。
整体来看,在陈酿6个月时,仅有籽单宁处理组可以观察到同对照组的颜色差异(ΔE > 3)。而对于陈酿12个月的酒样,皮单宁和籽单宁处理组均可观察到与对照组的颜色差异,这一现象可能是由酒样中花色苷类物质的含量引起的。
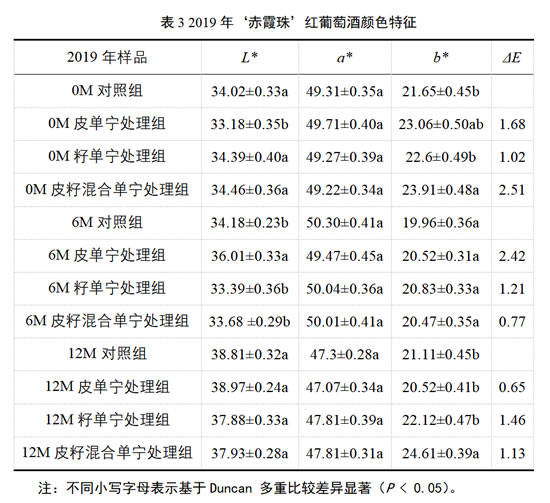
由表3可知,对2019年的‘赤霞珠’葡萄酒而言,经过6个月的桶储陈酿后,皮单宁处理组酒样L*值显著高于其他处理组,但当陈酿进行到12个月时,各处理组葡萄酒的L*值无显著差异。除此之外,各处理组的a*值在不同的陈酿取样点处,并未表现出显著的差异。而对于各处理组的b*值,在陈酿12个月时,只有添加了葡萄皮籽混合单宁的处理组最高,且显著高于其他处理组。
系统来看,2019年‘赤霞珠’葡萄酒在添加了不同来源的商业单宁后,在陈酿的各个时间点,各处理组与对照组之间的色差值ΔE均小于3,这表明各处理组同对照组之间不存在肉眼可以识别的颜色差异。
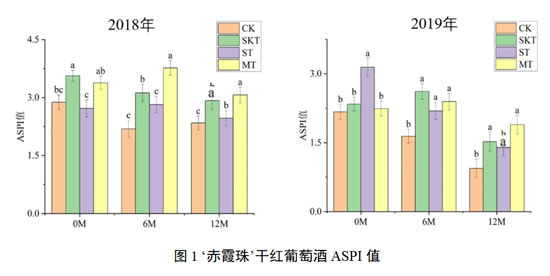
对于‘赤霞珠’葡萄酒的涩感品质而言,如图1所示,添加了不同来源的葡萄单宁后,2018年各处理组酒样的涩感强度均显著高于空白对照组,其中皮籽混合单宁处理组的效果最好。刚入桶时,皮单宁与皮籽混合单宁均能迅速提高‘赤霞珠’葡萄酒的涩感强度,且这种效果在桶储过程中(6M和12M)均能得到有效保持,籽单宁的作用弱于皮单宁和皮籽混合单宁,但在陈酿后期仍显著高于对照组。2019年的‘赤霞珠’葡萄酒涩感强度表现与2018年类似,经历了12个月的桶储陈酿后,各处理组酒样的涩感强度均有显著提高,其中皮籽混合单宁组仍表现出最强的提升涩感的能力。与2018年实验结果不同的是,入桶后籽单宁组能迅速提升‘赤霞珠’葡萄酒的涩感强度,而其他处理组对涩感的影响与对照组无显著区别,且在陈酿6个月时,皮单宁和皮籽单宁处理组的涩感强度得到了显著提升。
3.2 不同来源葡萄单宁对陈酿过程中‘马瑟兰’葡萄酒颜色和涩感的影响
如表4所示,对于2018年的‘马瑟兰’葡萄酒,其L*、a*、b*值,在陈酿过程中,对照组及各处理组之间均不存在显著性差异。在陈酿12个月时,籽单宁处理组的酒体颜色最浅,其L*值显著高于对照组及其他处理组,且其b*值最低,这表明在添加葡萄籽单宁后,‘马瑟兰’酒样黄色色调减弱,葡萄籽单宁可能有助于延缓‘马瑟兰’干红葡萄酒的黄化。然而,综合对比色差值ΔE的数据可知,在整个陈酿过程中,对照组同各处理组之间均不存在肉眼可以辨识的颜色差异。
而对于2019年‘马瑟兰’葡萄酒(表5),在陈酿6个月时,皮籽混合单宁处理组的L*值最高,且显著高于对照组与其他处理组,同时其b*值最低,表明酒样具有最浅的颜色和最低的黄色色调。并且当陈酿时间达到12个月时,皮籽混合单宁处理组仍保持了最低的b*值,显著减弱了酒样的黄化程度。同18年‘马瑟兰’葡萄酒添加实验类似的是,在整个陈酿过程中,对照组同各处理组之间也均不存在肉眼可以辨识的颜色差异,其ΔE值均低于3。这一现象与以往研究存在一定差异,可能原因是‘马瑟兰’是一种富含花色苷类物质的酿酒葡萄品种,其所酿造的葡萄酒产品颜色表现较好,因此添加单宁等辅料后,对酒体本身的影响是有限的。除此之外,也可能是陈酿时间较短,难以明确添加不同来源葡萄单宁对‘马瑟兰’葡萄酒颜色稳定性的影响。
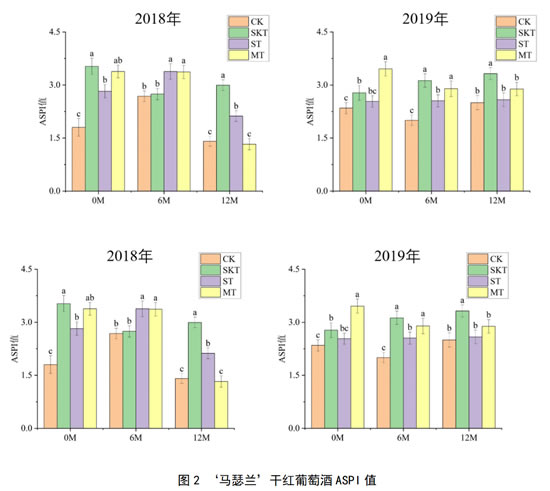
图2展示了两个年份‘马瑟兰’葡萄酒在桶储陈酿过程中的涩感强度变化,由图可知,2018年‘马瑟兰’葡萄酒在添加了不同来源的葡萄单宁后,各处理组的涩感强度在不同的陈酿时间表现出了一定的差异:在陈酿6个月时,籽单宁处理组和皮籽混合单宁处理组的涩感强度显著高于皮单宁处理组,而在陈酿12个月后,葡萄皮单宁对‘马瑟兰’葡萄酒涩感强度的作用更大,其ASPI结果显著高于其他处理组。葡萄籽单宁的添加,有助于陈酿过程中酒样涩感强度的提升与稳定,但这种效果更多的体现在陈酿中期,随着陈酿时间的进行,葡萄皮单宁的作用更佳。
2019年‘马瑟兰’葡萄酒在刚入桶时,皮单宁和皮籽单宁能快速提升酒体涩感强度,且这种作用效果可一直持续至陈酿12个月,籽单宁的加入虽然也能适当增强‘马瑟兰’葡萄酒的涩感强度,但这种作用似乎只能维持到陈酿中期(6M),当陈酿时间达到1年时,籽单宁处理组的涩感强度与对照组无显著性差别。综合2年的结果可以发现,添加皮单宁对‘马瑟兰’葡萄酒的提升效果更为显著以及稳定。
3.3 不同来源葡萄单宁对陈酿过程中‘赤霞珠’葡萄酒酚类物质衍变影响
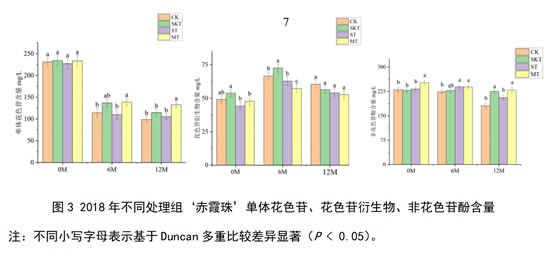
2018年的‘赤霞珠’葡萄酒的单体花色苷、花色苷衍生物和非花色苷酚含量如图3所示:在刚入桶时,添加不同来源的葡萄单宁对酒样中的单体花色苷含量无显著影响,而葡萄皮单宁的添加可提升酒样中花色苷衍生物含量。在桶储陈酿6个月时,只有添加了葡萄皮单宁的处理组酒样中的花色苷衍生物显著高于对照组,添加了葡萄籽单宁和皮籽混合单宁的处理组中花色苷衍生物的含量均低于对照组。而经过了12个月的陈酿后,皮籽混合单宁处理组单体花色苷含量显著高于其他处理组和对照组,各处理组花色苷衍生物的含量均低于对照组,皮单宁处理组和皮籽混合单宁处理组非花色苷酚含量显著高于对照组。
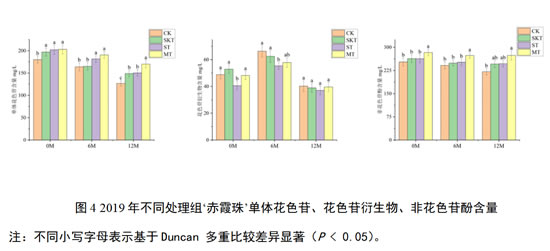
类似地对2019年‘赤霞珠’葡萄酒中单体花色苷、花色苷衍生物和非花色苷酚含量进行了分析:在刚入桶时,葡萄单宁的添加能够显著提升葡萄酒中的单体花色苷含量,这与2018年结果并不一致;葡萄籽单宁的加入显著降低了花色苷衍生物的含量;葡萄皮籽混合单宁的加入能够显著增加酒中非花色苷酚的含量。在经过12个月的陈酿后,添加了葡萄单宁的各处理组,其对单体花色苷和非花色苷酚的保护效果均好于对照组,其中添加了葡萄皮籽混合单宁的处理组对这些酚类物质的保护最为有效。但对于花色苷衍生物来说,添加不同来源的葡萄单宁在桶储陈酿12个月后不会影响其积累。
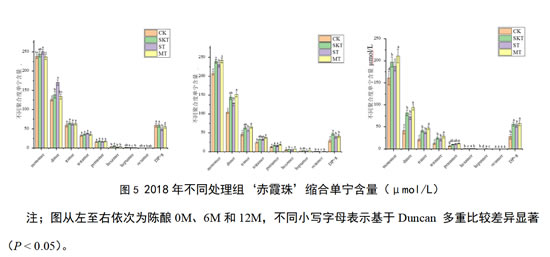
本研究进一步解析了桶储陈酿阶段,各处理组缩合单宁轮廓的演变情况。如图5所示,2018年‘赤霞珠’葡萄酒在刚添加各类单宁并入桶时,籽单宁处理组能够显著提高大部分不同聚合度的缩合单宁的含量,皮籽混合单宁处理组次之。在陈酿12个月后,皮籽混合单宁处理组其单体、二聚体、三聚体和四聚体含量显著高于对照组。
总的来说,对于2018年‘赤霞珠’葡萄酒中的黄烷醇单体的含量随陈酿进行而逐渐降低;对于酒中的低聚单宁,其含量主要呈现随陈酿时间逐渐降低的趋势,这可能是酒中高聚单宁逐渐分解,或是单体、二聚体不断聚合导致的,酒中的高聚单宁随陈酿时间增加而逐渐升高,但籽单宁处理组呈现随陈酿时间增加先降低后增加的趋势。
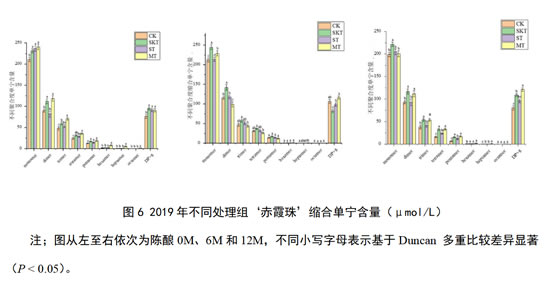
如图6所示,2019年‘赤霞珠’葡萄酒在刚入桶时,皮籽混合单宁处理组中单体、二聚体、三聚体含量显著高于对照组及其他处理组;添加了葡萄单宁的各个处理组,其聚合度大于8的缩合单宁含量均得到显著提升。在陈酿6个月时,皮单宁处理组中单体、二聚体、三聚体及四聚体和五聚体含量明显更高。而在陈酿12个月后,皮单宁处理组中单体、二聚体、三聚体含量显著高于对照组,得到了较好的保持,但对于聚合度大于8的缩合单宁,此外,添加皮籽混合单宁的处理组其除单体外各聚体单宁含量得到显著增加,对于聚合度大于8的单宁,皮籽混合单宁处理组含量显著高于对照组和其他处理组。
3.4 不同来源葡萄单宁对陈酿过程中‘马瑟兰’葡萄酒酚类物质衍变影响
对2018年‘马瑟兰’葡萄酒中单体花色苷、花色苷衍生物和非花色苷酚含量进行了分析(图7):在刚入桶时,对照组和添加不同来源葡萄单宁的处理组之间的单体花色苷和花色苷衍生物含量无显著差异;皮单宁处理组和皮籽混合单宁处理组显著增加了总非花色苷酚的含量。在陈酿12个月后,添加葡萄皮单宁或添加葡萄籽单宁进行陈酿均能使得花色苷衍生物和非花色苷酚的含量显著高于对照组,由上述现象可知,添加不同来源的葡萄单宁进行陈酿,其对于葡萄酒中的单体花色苷含量影响较小;添加单宁进行陈酿能够促进花色苷衍生物的形成,并减缓其降解,同时也能维持葡萄酒中非花色苷酚含量的稳定。
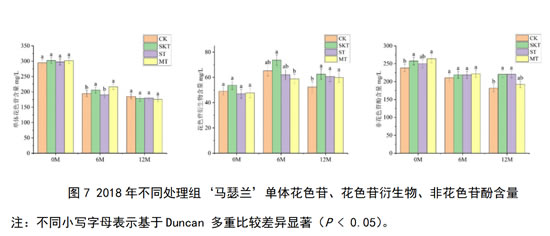
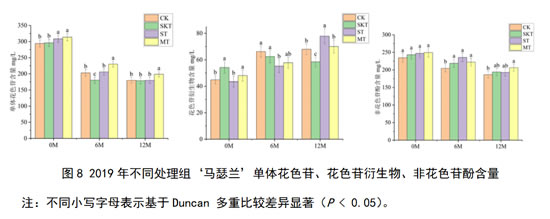
对于2019年‘马瑟兰’葡萄酒而言,其单体花色苷、花色苷衍生物和非花色苷酚含量如图8所示,在刚入桶时,葡萄籽单宁和皮籽混合单宁的添加能够显著提升葡萄酒中的单体花色苷的含量;葡萄皮单宁和皮籽混合单宁显著增加了酒中花色苷衍生物的含量。在经过12个月的陈酿后,添加了葡萄单宁的各处理组,其对单体花色苷和非花色苷酚的保护效果均好于对照组,其中添加了葡萄皮籽混合单宁的处理组对花色苷衍生物的保护最为有效。对于花色苷衍生物来说,添加不同来源的葡萄单宁在桶储陈酿12个月后均能显著增加其含量,促进花色苷衍生物积累。
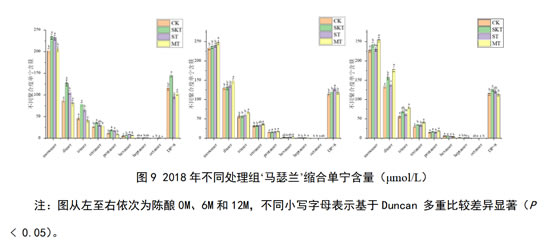
除上述酚类物质外,本研究进一步解析了桶储陈酿阶段,各处理组缩合单宁轮廓的演变情况。如图9所示,对于2018年‘马瑟兰’葡萄酒,在刚入桶时,添加不同来源的单宁均可有效的提高各个聚体的缩合单宁的含量。皮单宁处理组对于各个聚体的单宁含量提升最为显著。经12个月的陈酿后,皮籽混合单宁处理组能够显著增加大部分不同聚合度的单宁含量,而这一趋势在陈酿6个月时就有所显现。综上,对于未经陈酿的‘马瑟兰’葡萄酒,添加葡萄皮单宁能够快速提升不同聚合度缩合单宁的含量。但对于陈酿时间较长的酒样,葡萄皮籽混合单宁的添加更能够稳定维持酒样中各聚合度单宁的含量。
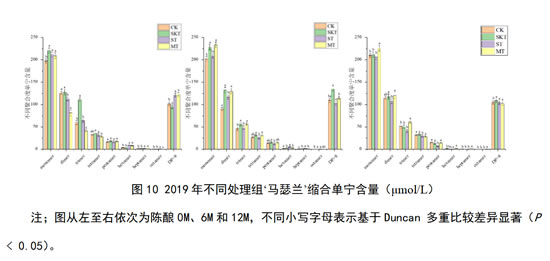
由图10,对于2019年‘马瑟兰’葡萄酒,对其对照组及不同处理组的缩合单宁含量进行分析:在刚入桶时,添加葡萄皮单宁可显著提高酒中大部分不同聚合度单宁含量,对于酒样中聚合度大于8的单宁来说,主要是葡萄籽单宁的作用使其含量提高。在经过12个月的陈酿后,能够提高酒样中各聚合度单宁含量的处理组是皮单宁和皮籽混合单宁的处理组,其中,皮籽混合单宁处理组效果更为显著。这一现象在陈酿6个月是,就有所体现,在陈酿6个月时,葡萄皮单宁的添加对于单宁含量的提升效果优于陈酿12个月。
3.5 不同基质组成对葡萄酒颜色和涩感的影响
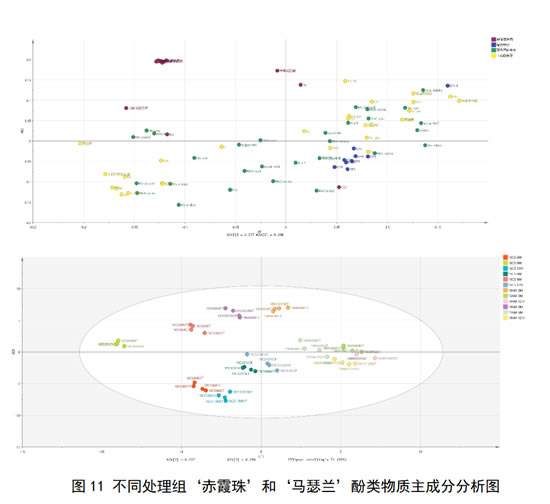
单宁的添加对于不同品种,不同年份、不同陈酿时间的葡萄酒,对其酚类物质演变和对于葡萄酒颜色和涩感的影响不尽相同,可能是由于不同葡萄酒本身不同的基质组成所带来的影响,故对于两个年份不同陈酿时间不同处理组的‘赤霞珠’和‘马瑟兰’葡萄酒的酚类物质进行主成分分析,由图11可知,首先,两个品种葡萄酒得到明显的区分,‘马瑟兰’葡萄酒主要分布在第一、四象限,和‘赤霞珠’葡萄酒得以明显区分。此外,对于两个品种葡萄酒进行单独分析,2018年刚入桶‘赤霞珠’酒样桶19年刚入桶酒样得以明显区分,表明两个年份赤霞珠酒样酚类物质轮廓有较大差异。18年刚入桶‘赤霞珠’酒样对应黄烷醇,表明其黄烷醇含量高,其距离高聚单宁较远,高聚单宁含量少,单宁加入后,能够有效维持单宁的含量,有效增强葡萄酒涩感,其花色苷衍生物含量少,单宁加入后有效促进花色苷衍生物生呈,使颜色有明显差异。2019年酒样对应花色苷衍生物,其高聚单宁含量多,富含花色苷衍生物,单宁加入后,对酒样本身单宁维持效果一般,对花色苷影响小,故酒样间未产生明显颜色差异。
对于‘马瑟兰’葡萄酒,其富含单体花色苷花色苷衍生物,单宁添加对其花色苷影响有限,对各个处理的颜色的表现无明显区别。马瑟兰富含高聚单宁,添加单宁陈酿后对其单宁含量影响有限,故对于涩感强度影响也有限。
3.6 结论
(1)不同来源葡萄单宁对葡萄酒的酚类物质组成具有重要影响。对于‘赤霞珠’葡萄酒,添加葡萄皮单宁在陈酿初期显著增加了花色苷衍生物和非花色苷酚的含量,并且能有效维持高聚单宁稳定性;葡萄籽单宁的添加能够增加其低聚单宁含量,但其效果不如在‘马瑟兰’葡萄酒中突出;葡萄皮籽混合单宁的添加在陈酿后期对单体花色苷和非酰化花色苷的保护作用最为突出。‘马瑟兰’葡萄酒添加葡萄籽单宁和葡萄皮籽混合单宁均能显著提高非酰化花色苷和乙酰化花色苷的含量,且葡萄籽单宁的加入可促进酒中低聚单宁的积累,葡萄皮籽混合单宁能有效改变酒样中低聚单宁含量。葡萄皮单宁的添加能快速增加黄烷醇和羟基苯甲酸的含量,也能促进‘马瑟兰’葡萄酒缩合单宁四聚体到七聚体形成。
(2)橡木桶陈酿前添加葡萄单宁葡萄酒的颜色和涩感影响存在差异。在葡萄酒颜色方面,对于‘赤霞珠’葡萄酒,葡萄籽单宁的加入显著增加了黄色色调,加速了酒液的黄化;对于‘马瑟兰’葡萄酒,葡萄籽单宁的添加明显降低了酒样的黄色色调,对酒液黄化有一定的延缓作用。葡萄皮单宁的加入能够显著提升‘赤霞珠’葡萄酒的花色苷衍生物含量,有助于增强红色色调;对于‘马瑟兰’葡萄酒,添加葡萄皮单宁有助于延缓酒液黄化。葡萄皮籽混合单宁的添加有助于保持‘马瑟兰’葡萄酒的红色色调,有助于稳定酒体颜色。在葡萄酒的涩感方面,对于‘赤霞珠’葡萄酒,葡萄皮单宁的添加显著提升了陈酿后的涩感强度,这一作用在陈酿后期更为突出。葡萄籽单宁的添加在陈酿初期显著增强了涩感,但在陈酿后期效果减弱,对于‘马瑟兰’葡萄酒,葡萄籽单宁的加入对涩感的提升效果优于‘赤霞珠’。葡萄皮籽混合单宁的添加在两种葡萄酒中均表现出较好的涩感提升效果,尤其是在陈酿中期效果明显。对于葡萄酒的单宁而言,葡萄皮单宁的加入更有效地维持‘赤霞珠’中高聚单宁的稳定性。
(3)‘赤霞珠‘葡萄酒花色苷含量中等,非花色苷酚类尤其黄烷醇和羟基肉桂酸含量高,缩合单宁含量较高,陈酿过程中高聚单宁占比增加,适合使用葡萄皮单宁或者葡萄皮籽混合单宁增强涩感和颜色稳定性,提高酒中高聚单宁、花色苷衍生物及黄烷醇等具有护色作用的非花色苷酚物质的含量。‘马瑟兰’葡萄酒花色苷含量丰富,非花色苷酚如羟基苯甲酸含量较高,缩合单宁含量较低,使用葡萄单宁改善颜色的效果有限,使用葡萄籽单宁或葡萄皮籽混合单宁能有效提升涩感,提高低聚单宁和羟基苯甲酸的含量。