元谋综合试验站
饶余倩 黄亚倩 夏龙腾 杨国顺 王美军 许延帅 刘昆玉
摘要:【目的】为寻找‘阳光玫瑰’葡萄理想安全的破眠方法以及探究其破眠机理。【方法】本研究以‘阳光玫瑰’葡萄冬芽为试材,通过研究类休眠阶段和生理休眠阶段不同破眠处理对葡萄萌芽率、成花率和花穗质量的影响,并通过转录组测序筛选了可能影响葡萄冬芽萌发的关键基因。【结果】在萌芽前1个月对‘阳光玫瑰’葡萄冬芽进行单氰胺处理、石硫合剂+ACC处理,均能提前使萌芽提前,显著提高萌芽率,不影响成花率及花穗质量。通过分析转录组数据,筛选了PYL4、PP2C24、PP2C8、PP2C37、PP2C25、SAPK2等差异表达基因,这些基因可能通过参与激素信号传导、氧化应激等途径影响葡萄芽的萌发。【结论】石硫合剂+ACC的混合溶液是‘阳光玫瑰’葡萄打破休眠的一种安全、有效的药剂。
关键词:葡萄;休眠;萌芽;转录组
葡萄(Vitis vinifera. spp)是多年生落叶藤本植物,栽培历史悠久。中国是葡萄生产大国,2020年我国葡萄产量为1431.4万吨,居世界前列。休眠是落叶果树的典型特征,是植物应对外界环境变化和季节变化的一种生物适应性,是植物生长发育过程中的一个周期性时期。葡萄作为一种典型的落叶果树,萌芽整齐度与产量密切相关。南方地区是近年来发展最快的葡萄产区之一,但在南方地区,由于树形培养管理(一年生枝条的培养)不当和需冷量积累不足,导致萌芽不整齐,造成在后期冬季修剪和盛花期花果管理上更加费时费工等问题。目前,单氰胺虽被广泛应用于打破葡萄休眠,但其本身具有很强的毒性,使用过程中需要戴口罩、手套等,操作不便。因此,寻找一种可以有效打破休眠且理想安全的破眠方法尤为重要。
多年生落叶植物芽休眠是一种十分复杂的过程,受到多条通路基因的综合调控。当植物芽开始进入休眠阶段时,可以抵御外界不良条件,暂时停止生长,当外界环境适宜时,植物可恢复正常生长,是一种对植物有利的生物学特征。1987年,Lang等根据休眠的诱因把芽休眠分为内休眠、类休眠和生态休眠三大类,并将休眠的整个过程分为五个阶段:类休眠、类休眠-内休眠、内休眠、内休眠-生态休眠和生态休眠[1]。内休眠(生理休眠,Endodormancy)发生在冬季,由植物内部原发性反应引起的生长停滞现象,这种原发性反应由休眠组织内部信号独自诱导引发。处于内休眠阶段的芽,即使在有利的环境条件,且没有附近器官的限制,也不能萌动,只有达到一定的低温积累后才能解除休眠。类休眠(Paradormancy)是由分生组织外的因子引起的植物生长暂停,本质上是一个器官对另一个器官的影响,如芽的顶端优势。处于类休眠阶段的芽,即使在环境条件有利时仍保持休眠,但若除去相邻器官的限制源,则休眠结构会迅速恢复生长。生态休眠(Ecodormancy)是指冬芽从生理休眠中释放后,由于环境因子限制暂未萌发,环境条件适宜时即可迅速恢复生长。
休眠和萌发是两个连续的生物过程,芽休眠是温带落叶果树的一种适应机制。葡萄的内休眠需要在经历一定的低温积累后才能自然打破,这种特性成为“需冷量”(Chilling requiremengt),葡萄的需冷量一般需要0~7.2℃的低温积累1000 h左右。为使落叶果树能够在低纬度地区正常生长发育,人们通常通过使用氰氨类、硝酸盐类、植物生长调节剂(GA3、6-BA、TDZ等)、大蒜汁、乙烯、破眠剂1号等化学物质来代替部分低温的作用,提早打破休眠,使休眠芽提前萌发。
植物激素在芽休眠的形成与解除过程中发挥重要的调节作用,不同的植物激素以不同的作用方式来诱导和解除芽休眠。ABA对萌芽有显著的抑制作用。在梨、葡萄、桃和蓝莓等多年生落叶果树中,外源ABA可以促进芽的休眠而抑制芽的萌发。在桃花芽的休眠过程中,ABA的含量在生态休眠期逐渐降低并达到最低水平。杨博等研究表明由PpyERF060、PpyABF3和PpyMADS71构成的互作网络可整合乙烯与脱落酸的信号通路进而调控梨芽休眠进程。玉米分蘖芽发生过程中,叶片与基部茎节中ABA含量较低且呈下降的趋势,当分生长停止时ABA含量上升。在杨树上,转录因子SVL可以通过促进CALS1(胼胝质合酶基因)的表达,抑制赤霉素的合成,控制自然休眠。在桃中,外施GA可以促进桃叶内休眠的解除。梨花芽可能通过下调miR159表达,促进GAMYB提高GA含量促进花芽冬季休眠的解除。乙烯对葡萄芽休眠解除起一定的促进作用。在葡萄的研究中发现外源乙烯的处理使葡萄芽的萌发率显著提高,而抑制乙烯合成和信号转导后,葡萄芽的萌发受到抑制,推测乙烯可以作为刺激信号促进葡萄芽休眠提前解除。Khalil-Ur-Rehman研究结果ABA、乙烯、赤霉素、生长素和细胞分裂素均参与了葡萄芽类休眠和内休眠的调控。
本研究对处于两种冬芽休眠阶段(类休眠阶段和内休眠阶段)的‘阳光玫瑰’葡萄冬芽进行不同破眠处理,以未处理作为对照,进行单氰胺涂芽处理和石硫合剂+ACC涂芽处理。记录和观察处理后‘阳光玫瑰’葡萄冬芽的生长发育动态,并统计其萌芽率、成花率及花穗质量。通过采用基于RNA-seq分析,筛选不同休眠阶段‘阳光玫瑰’葡萄冬芽破眠的相关基因表达情况,通过对差异表达基因进行KEGG代谢通路分析探索不同休眠阶段‘阳光玫瑰’葡萄冬芽休眠解除的关键影响基因。
1 材料与方法
1.1 试验材料
试验于2022-2023年在湖南农业大学教学基地内进行,供试品种为四年生‘阳光玫瑰’葡萄。栽培方式为起高垄式根域限制栽培,喷灌方式为微喷。试验所用试剂:石硫合剂(有效成分含量29%,山东东信生物农药有限公司);单氰胺(有效成分含量50%,宁夏大荣化工冶金有限公司);1-氨基环丙烷羧酸(ACC,纯度98%,上海源叶生物科技有限公司)。
1.2 试验方法
1.2.1 冬季(内休眠阶段)冬芽破眠处理与样品采集
2021年1月,对‘阳光玫瑰’基部留2芽修剪,收集基部起第3节位以上的枝条78根。于2022年2月28日选取3~9节位(9个冬芽)的枝条进行破眠试验,各处理如下,每个处理26根:

分别在0 h、处理后24 h和72 h收集冬芽,样品收集标准:每个处理随机取4根枝条,收集每根枝条全部的冬芽,并迅速放入液氮中速冻,随后保存在-80 ℃超低温冰箱备用。
2023年2月,在六年生“飞鸟”架活体‘阳光玫瑰’葡萄树上重复上述试验验证结果。
1.2.2 夏季(类休眠阶段)冬芽破眠处理与样品采集
2022年8月13日,将平棚架“一”字型‘阳光玫瑰’采收后的一年生主梢进行摘叶处理,将主梢进行第7节位短截后对其第5-7节位上的冬芽进行不同破眠剂处理(处理方法同表1),每个处理53根枝条。分别在0 h、处理后24 h和72 h收集冬芽,样品收集标准:每个处理随机取10根枝条,每根枝条上取5-7节位上的3个芽,并迅速放入液氮中速冻,随后保存在-80 ℃超低温冰箱备用。
1.2.3 观测指标与方法
以花芽露绿50%作为萌发指标,进行冬芽萌芽率的统计。
萌芽率=已萌动的冬芽数/调查的冬芽数
成花率=有花穗新梢数/已萌动冬芽数
1.2.4 RNA提取、文库构建和转录组测序
将2022年2月和8月收集的冬芽样品的RNA提取以及转录组测序委托百迈客生物科技有限公司(北京)完成。使用Nanodrop2000(赛默飞,Nanodrop2000)对于提取的RNA进行浓度检测,并使用Agient2100,LabChip GX(厂家铂,型号铂铂金埃尔默LabChip GX)对完整性进行检测。样品检测合格后,进行文库构建:用带有Oligo(dT)的磁珠富集真核生物mRNA 加入Fragmentation Buffer将mRNA进行随机打断;以mRNA为模板,合成第一条cDNA链及二链,并进行cDNA纯化;纯化的双链cDNA再进行末端修复、加A尾并连接测序接头,然后用AMPure XP beads进行片段大小选择;最后通过PCR富集得到cDNA文库。经库检合格后,使用Illumina NovaSeq6000测序平台进行PE150模式测序。
1.2.5 差异表达基因的筛选和分析
首先对样品中的Mapped Reads的数目和转录本长度进行归一化,以FPKM方法作为检测转录本或基因表达水平的指标。不同样本中表达水平存在显著差异的基因称之为差异表达基因(DEGs)。根据不同样品之间表达水平的相对高低,差异表达基因可以分为上调基因(Up-regulated Gene)和下调基因(Down-regulated Gene)。使用edgeR软件进行差异分析,将|log2Fold Change|≥1且FDR<0.01作为差异表达基因的筛选标准。使用百迈客分析平台导出基因在不同样品中的功能注释和相关代谢通路等信息,差异表达基因基于GO数据库和KEGG数据库进行功能注释并按照生物学过程、细胞组分和分子功能对其富集分析,找出差异基因显著富集的GO term和KEGG term。
1.2.6 数据分析
所有数据的采集和计算均设3组生物学重复。运用Excel整理数据,GraphPad Prism8和R语言聚类程序作图;SPSS26.0进行单因素ANOVA分析,多重比较采用LSD法。
2 结果与分析
2.1 不同破眠处理对不同休眠阶段葡萄萌芽和开花的影响
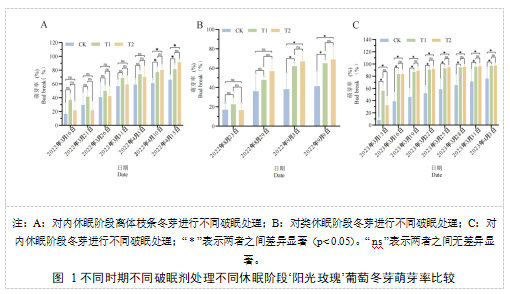
不同破眠剂处理会对不同休眠阶段的‘阳光玫瑰’葡萄芽的萌芽率、萌芽速度均会产生一定影响(图1)。内休眠阶段葡萄离体枝条冬芽萌芽率结果(图1-A)显示,T1处理萌芽最早,其次是T2处理,最后是CK组,各处理萌芽整体速度呈现先慢后快的趋势(图1-A)。2022年4月10日T1、T2处理后的萌芽率与CK相比开始出现差异显著,其中T2处理的萌芽率最高。2022年4月15日T1处理与T2处理的萌芽率分别为81.48%和85.80%,显著高于CK组(66.05%)。类休眠阶段葡萄冬芽萌芽率结果(图1-B)显示,T1处理萌芽最早,2022年9月3日T1、T2处理后的萌芽率与CK相比开始出现差异显著,其中T2处理的萌芽率最高。2022年9月9日T1处理与T2处理的萌芽率分别为65.66%和69.70%,显著高于CK组(41.41%)。内休眠阶段葡萄冬芽萌芽率结果(图1-C)显示,处理后25天T1、T2处理后的萌芽率与CK相比开始出现差异显著,2023年4月3日T1处理与T2处理的萌芽率分别为96.43%和97.22%,显著高于CK组(76.19%)。
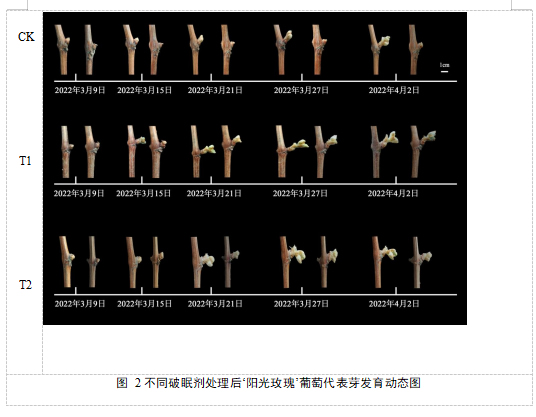
破眠处理后内休眠阶段离体枝条上的葡萄芽的生长发育动态如图2所示,T1、T2处理的葡萄芽提早解除休眠,均在2022年3月21日出现2-3枚新叶,整体呈现先慢后快的趋势。T1处理在2022年3月15日已经开始萌芽,萌芽开始时间最早,萌芽较为整齐;其次是T2处理的葡萄芽在3月21日左右完成萌芽期,萌芽较为整齐。CK组的葡萄芽在2022年4月2日出现2-3枚新叶,生长相对缓慢,萌芽不整齐。
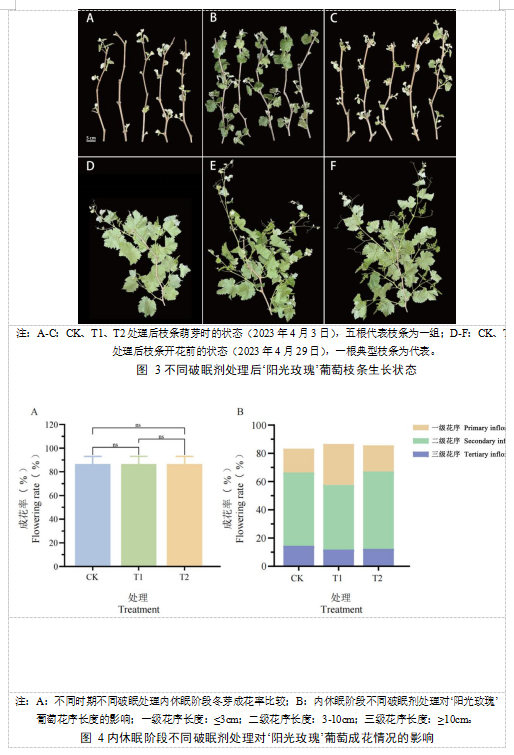
破眠处理后内休眠阶段葡萄枝条生长状态如图3所示,相比于CK组,2023年4月3日T1处理后的叶片生长速度最快,其次是T2处理。2023年4月29日T1、T2处理后的叶片生长速度、叶片大小、节间长度与CK组相比无显著影响。
为了解内休眠阶段不同破眠处理对葡萄花穗的影响,本研究统计了不同破眠处理后所有一年生枝条的成花情况。图4显示,内休眠阶段T1、T2处理对葡萄一年生枝条的成花率相比于CK组没有出现差异显著,同时也不影响花序长度。
2.2 转录组测序数据质量分析及序列比对
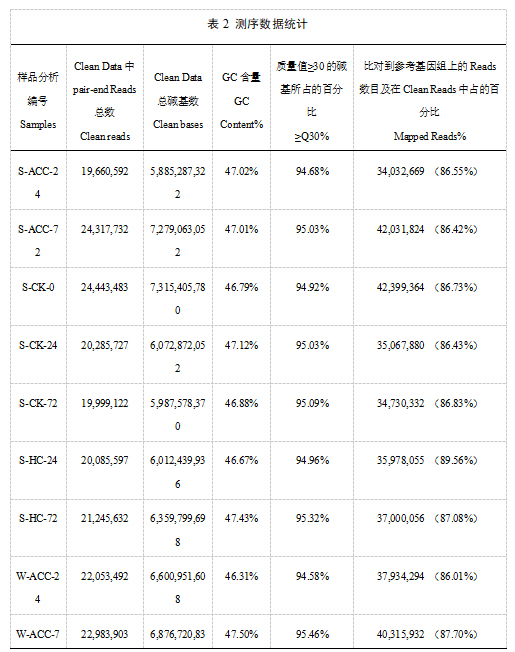
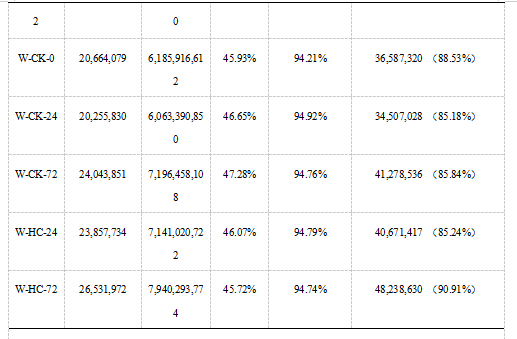
利用Illuminate平台,完成14个样本的转录组测序。为保证后续分析的准确,我们通过对原始数据进行一系列严格的质量控制后得到高质量的Clean Data。经过测序质量控制后,共得到92.92Gb Clean Data,各样品Clean Data均达到5.89Gb,Q30碱基百分比在94.21%及以上,GC含量为45.72%~47.50%。表明测序质量比较好,可以用于后续的比对分析。通过使用HISAT2将Clean Reads与参考基因组进行比对后发现,各样本的Reads与指定的参考基因组的比对率在85.18%~90.91%。
2.3 差异表达基因的筛选
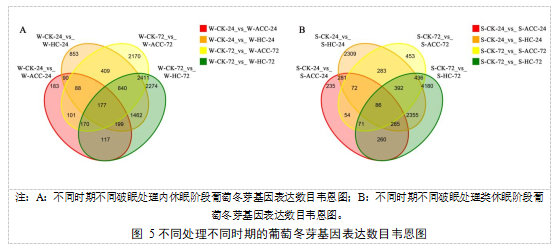
以|log2fold changes|≥1、FDR<0.01为筛选条件。通过总差异韦恩图(图5)分析发现,在内休眠阶段,6个处理组中全部共有的差异基因为177个,W-CK-72中的样品与W-ACC-72中的样品比和W-HC-72中的样品比有大量相同的差异基因,为2411个;W-CK-24中的样品与W-ACC-24中的样品比和W-CK-72中的样品与W-ACC-72中的样品比共有差异基因101个;W-CK-24中的样品与W-HC-24中的样品比和W-CK-72中的样品与W-HC-72中的样品比共有差异基因1462个;W-CK-24中的样品与W-ACC-24中的样品比和W-HC-24中的样品比差异基因较少,仅90个。在类休眠阶段,6个处理组中全部共有的差异基因为86个,S-CK-24中的样品与S-HC-24中的样品比和S-CK-72中的样品与S-HC-72中的样品比有大量相同的差异基因,为2355个;S-CK-72中的样品与S-ACC-72中的样品比和S-HC-72中的样品比共有差异基因436个;S-CK-24中的样品与S-ACC-24中的样品比和S-HC-24中的样品比共有差异基因281个;S-CK-24中的样品与S-ACC-24中的样品比和S-CK-72中的样品与S-ACC-72中的样品比共差异基因较少,仅54个。
2.4 休眠期间差异表达基因模式变化
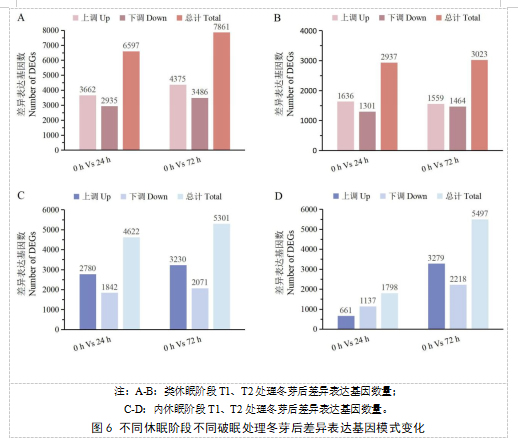
研究了不同休眠阶段不同破眠剂处理后葡萄芽2个时期的基因表达模式,不同时期样本之间的DEGs数量被鉴定(图6).结果表明,类休眠阶段,T1处理后24小时有3662个基因显著上调,2935个基因显著下调;T1处理后72小时有4375个基因显著上调,3486个基因显著下调;T2处理后24小时有1636个基因显著上调,1301个基因显著下调;T2处理后72小时有1559个基因显著上调,1464个基因显著下调。内休眠阶段,T1处理后24小时有2780个基因显著上调,1842个基因显著下调;T1处理后72小时有3230个基因显著上调,2071个基因显著下调;T2处理后24小时有661个基因显著上调,1137个基因显著下调;T2处理后72小时有3279个基因显著上调,2218个基因显著下调。
2.5 休眠期间差异基因的聚类分析
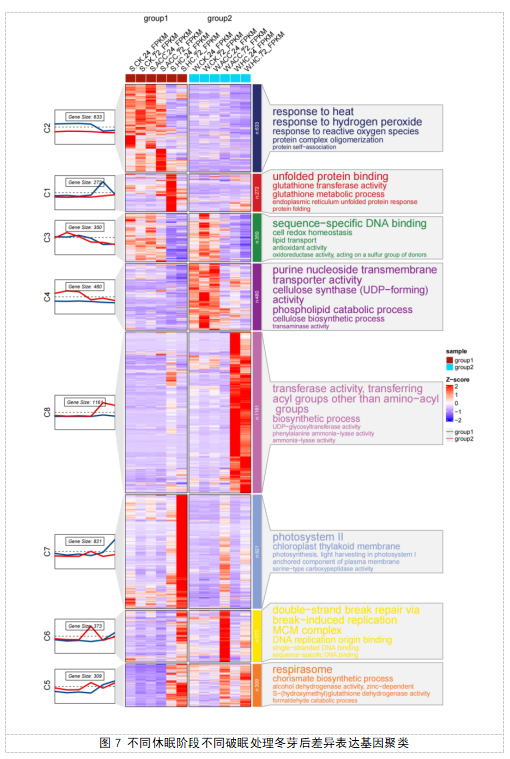
表达模式相似的基因通常具有相似的功能,为了理解不同休眠阶段不同破眠处理后不同时期芽的差异表达基因的表达模式,我们利用R语言中的聚类程序对12组样本的显著差异基因进行等级聚类分析(图7)。结果表明,4399个基因被分成8组,最多的一组(Class 8)包含了1161个DEGs,DEGs的表达模式呈现为在W-HC-24样本、W-HC-72样本上调表达,其他样本基本下调表达,这些基因显著富集在转移酶活性,转移氨基酰基以外的酰基、生物合成进程、UPD-糖基转移酶活性、苯丙氨酸解氨酶活性、解氨酶活性等途径。
第二大组(Class 7)包含821个基因,DEGs的表达模式呈现为在S-HC-72样本上调表达,其他样本基本下调表达,这些基因主要富集在光系统II、叶绿体类囊体膜、光合作用,光系统I的捕光天线复合体、膜的锚固成分、丝氨酸型羧肽酶活性等途径。
第三大组(Class 2)包含633个基因,DEGs的表达模式呈现为在S-CK-24样本、S-CK-72样本、S-ACC-24样本、S-ACC-72样本上调表达,其他样本基本下调表达,这些基因主要富集在响应热休克、响应过氧化氢、响应活性氧、蛋白质复合物寡聚化、蛋白质自聚集等途径。
第四大组(Class 4)包含480个基因,DEGs的表达模式呈现为在W-CK-24样本、W-CK-72样本、W-ACC-24、样本上调表达,其他样本基本下调表达,这些基因主要富集在嘌呤核苷跨膜转运蛋白活性、纤维素合酶 (UDP−forming)活性、磷脂代谢过程、纤维素生物合成过程、转氨酶活性等途径。
在Class 6中,373个基因在S-HC-72样本、W-ACC-72样本中上调表达,其他样本基本下调表达,这些基因主要富集在通过断裂诱导复制进行DNA双链断裂修复、微小染色体维持蛋白复合物、DNA复制起点结合、单链DNA结合、序列特异性DNA结合等途径。
在Class 3中,350个基因在S-CK-24样本、S-CK-72样本、S-ACC-24样本、S-ACC-72样本、W-CK-24样本、W-CK-72样本、W-ACC-24样本上调表达,其他样本基本下调表达,这些基因主要富集在序列特异性DNA结合、细胞氧化还原稳态、脂质转运、抗氧化活性、氧化还原酶活性、作用于硫基供体等途径。
在Class 5中,309个基因在S-HC-24样本、S-HC-72样本、W-ACC-72样本、W-HC-72样本上调表达,其余样本基因下调表达,这些基因主要富集在呼吸链、分枝酸生物合成过程、锌离子依赖的乙醇脱氢酶活性、S-羟甲基谷胱甘肽脱氢酶活性、甲醛分解代谢过程等途径。
在Class 1中,272个基因在S-HC-24样本中上调表达,其余样本基本下调表达,这些基因主要富集在未折叠蛋白反应、谷胱甘肽转移酶活性、谷胱甘肽代谢过程、内质网未折叠蛋白反应、蛋白质折叠等途径。
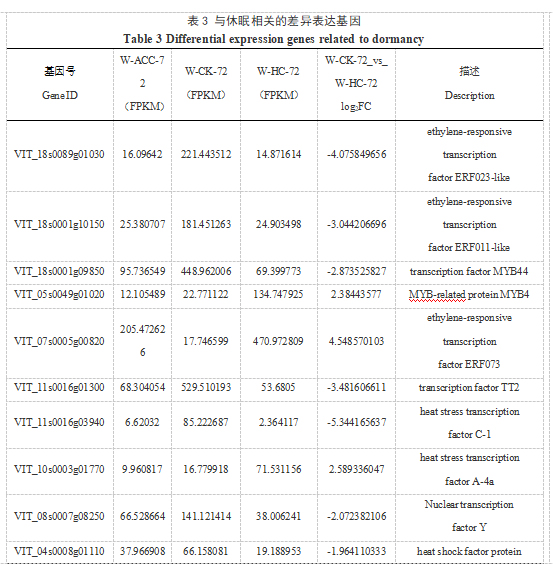
2.6 与休眠相关的差异表达基因筛选
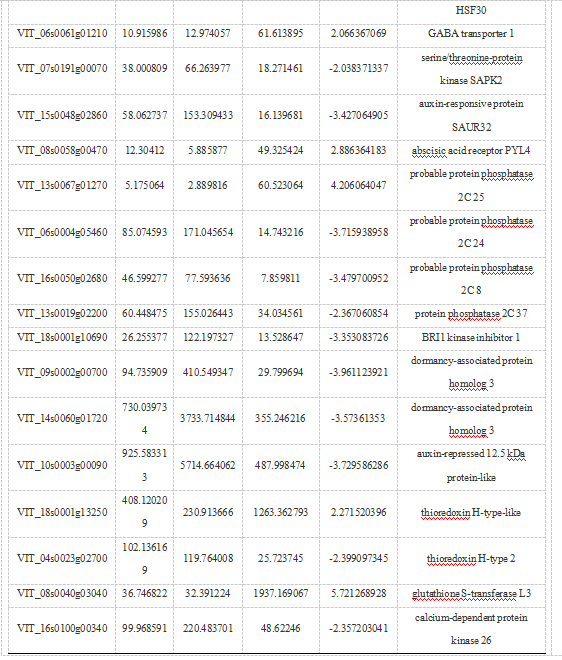
在使用不同破眠剂打破葡萄芽休眠的过程中会引起植物体内一系列生理生化变化,同时会诱导破眠相关基因的表达。在内休眠阶段W-CK-72样本与W-HC-72样本比的差异表达基因(表3)中有10被注释为转录因子编码基因,其中上调的转录因子有3个,包括MYB4、ERF073、HSF A-4a;下调的转录因子有7个,包括ERF023-like、ERF011-like、TT2、MYB44、HSF C-1、Nuclear transcription factor Y、HSF30。另外,有10个被注释为参与生长素(Auxin, AUX)、赤霉素(Gibberelli acid, GA)、脱落酸(Abscisic acid, ABA)、油菜素甾醇(Brassinosteroids, BR)等激素的生物合成或介导的信号转导过程。在ABA信号通路中,PYL4基因上调表达;4个与蛋白磷酸酶2C(PP2C)家族相关的基因被注释,PP2C24、PP2C8、PP2C37均下调表达,PP2C25上调表达;与Snrk2家族相关的SAPK2基因下调表达。剩下的差异基因与氧化应激、谷胱甘肽S-转移酶等有关。
3 讨论
生长期和休眠期是葡萄生长发育过程中较为明显的两个时期。在浅内休眠阶段,通过对葡萄冬芽进行物理或化学等破眠措施可使冬芽提前萌发。前人在不同品种葡萄(如‘巨峰’葡萄、‘户太八号’、‘香悦’葡萄等)上均已试验证明单氰胺可以提前打破葡萄休眠,在我国南方地区,萌芽前一个月对葡萄冬芽进行单氰胺处理破眠效果最佳,可明显提高萌芽率,从而在一定程度上提高能够提升葡萄产量。乙烯是五大植物激素之一,广泛参与到植物生长发育的各个进程中,前人研究证明植物器官本身能生成乙烯,并提出了乙烯具有促进植物成熟衰老的作用。乙烯是气体,在实际生产中主要使用乙烯利对香蕉、猕猴桃、油梨、杏等进行催熟,浙江、上海地区常用乙烯利对葡萄进行催熟。ACC是乙烯前体,在1-氨基环丙烷-1-羧酸氧化酶(ACC氧化酶,ACO)的氧化作用下生成乙烯。因此,我们推测ACC可能芽的萌发有一定的促进作用。
Keilin等在构建HC诱导表达的EST文库后,利用比较基因组学发现氧化应激、钙信号传导等生化过程中的功能基因可能参与休眠解除过程。Halaly等研究表明通过比较HC和热击(Heaat shock,HS)两种方式均可解除生理休眠,触发细胞生化水平暂时性呼吸和氧化应激,诱导CAT、ADH和PDC等相关基因瞬时上调表达,破眠处理还诱导抗坏血酸过氧化物酶、谷胱甘肽还原酶、硫氧还蛋白h、谷胱甘肽S-转移酶和蔗糖合成酶编码基因表达。单氰胺处理休眠芽后,通过释放氰化物抑制有氧呼吸和过氧化氢酶活性,导致细胞缺氧同时诱导过氧化氢产生,进而导致植物体内活性氧含量升高。Pacey-Miller等报道了葡萄休眠芽中活性氧相关的基因Cu/Zn超氧化物歧化酶、谷胱甘肽过氧化物酶、谷胱甘肽硫转移酶、谷胱甘肽还原酶和过氧化氢酶基因表达搞。在本研究中,MYB、ERF、MADS-box等转录因子都与芽萌发有关,这些转录因子通过共同调控多种途径从而影响葡萄芽萌芽,一些与氧化应激相关、钙信号传导等基因也从不同功能对葡萄活性氧代谢进行调控。
植物激素与芽休眠密切相关,尤其是ABA和GA在调控芽休眠和萌发的过程中发挥着重要作用。Khalil-Ur-Rehman等认为ABA和GA信号通路作为调节开关调节葡萄芽休眠。ABA可以促进杨树SVL的表达,而内源ABA可通过激活PpyABF3调控梨PpyDAM3的转录,从而调控其休眠进程。在梨芽内休眠解除后观察到较低水平的NCEDs表达,PpyCYB8和Ppy707A3的表达水平增加,同时ABA含量降低(Yang,2021)。李可等研究表明外源ABA可抑制蓝莓花芽萌芽。在本研究中,鉴定出与激素相关的基因,如GABA转运蛋白1、生长素反应蛋白、生长素抑制蛋白、脱落酸受体、BRI1激酶抑制剂1、蛋白磷酸酶、丝氨酸/苏氨酸蛋白激酶等可能均参与了植物激素信号转导。
本试验对不同休眠阶段的‘阳光玫瑰’葡萄冬芽进行破眠试验。用萌芽率、成花率及花穗质量作为衡量葡萄破眠效果优劣的三个指标,试验结果表明,在类休眠阶段和内休眠阶段,单氰胺处理、石硫合剂+ACC处理均对‘阳光玫瑰’葡萄萌芽产生积极影响,石硫合剂+ACC处理后的萌芽率略高于单氰胺处理,两种处理均能使‘阳光玫瑰’葡萄冬芽萌发时间提前,显著提升植株萌芽率,对后续成花率及花穗长度均无影响,且不会引起药害。在本研究中,还有一些与环境响应相关的差异基因,如热击蛋白(Heat shock protein,HSP)基因、光敏色素(phytochrome)基因等、水分代谢相关的基因,如水孔蛋白(aquaporin,AQP)基因等)、能量与物质代谢相关的基因,如蔗糖合酶(sucrose synthase)等、细胞发育相关的基因,如细胞周期蛋白(cyclin,CYC)基因、细胞周期蛋白依赖激酶(cyclin-dependent protein kinase,CDK)基因等)可能也参与了葡萄芽休眠解除的调控。
4.结论
‘阳光玫瑰’葡萄在内休眠、类休眠阶段经单氰胺和石硫合剂+ACC处理后,有提前萌芽的作用,且不影响后续成花率及花穗质量,对操作人员较为安全,打破葡萄休眠效果较好,石硫合剂+ACC处理萌芽率略高于单氰胺处理。石硫合剂+ACC的混合溶液是‘阳光玫瑰’葡萄打破休眠的一种安全、理想的药剂。单氰胺处理、石硫合剂+ACC处理均诱导了PYL4、PP2C24、PP2C8、PP2C37、PP2C25、SAPK2等基因的表达,这些基因可能参与了打破芽的休眠,促进萌芽。