苗木生产岗位
史宁 何非 王军
摘 要:砧木具有很强的抗生物和非生物胁迫能力,因此被广泛应用于葡萄栽培。在本研究中,我们进行了为期两年的研究(2016-2017年),采用高效液相色谱-质谱联用评估了5种砧木(101-14、1103P、Beta、5BB和SO4)对‘小味儿多’果实类黄酮的影响。结果显示,砧木对‘小味儿多’成熟速度的影响有限。在类黄酮化合物方面,1103P 和 Beta 连续两年都有增加葡萄皮花色苷含量的趋势。101-14 和 1103P 提高了果实黄酮醇的含量,而 Beta 则对黄酮醇的生物合成(丁香亭除外)产生了不利影响。此外,尽管年份因素对黄烷-3-醇的影响占主导地位,但 101-14 往往会使果皮和种子中的黄烷-3-醇浓度略有增加。总之,基于成熟葡萄浆果理化指标和类黄酮化合物的测定,本研究为中国东部地区‘小味儿多’的嫁接栽培提供了理论依据。
关键词:‘小味儿多’;砧木;嫁接;类黄酮
Modifications of Vitis vinifera L. cv. Petit Verdot grape flavonoids as affected by the different rootstocks in eastern China
Abstract: Rootstocks are widely used in viticulture because of their strong ability to resist biotic and abiotic stress. In two-successive years (2016-2017), the effects of five rootstocks (101-14, 1103P, Beta, 5BB, and SO4) on the berry flavonoids profiles of Petit Verdot were evaluated by HPLC-MS. The results showed that there was limited influence of rootstocks affecting the ripening of Petit Verdot. With respect to flavonoid compounds, 1103P and Beta tended to increase the concentrations of anthocyanin in grape skins in two seasons. 101-14 and 1103P enhanced the flavonol concentrations, whereas Beta showed a detrimental effect on the biosynthesis of flavonols except for the syringetin. Additionally, despite the season factor dominating the effect on flavan-3-ols, 101-14 tended to show a slight increase in flavan-3-ol concentrations in skins and seeds. In conclusion, based on the evaluation of physicochemical indicators and flavonoid compounds of mature berries, the current study provided a theoretical basis for grafting cultivation of Petit Verdot in eastern China. Keywords: Petit Verdot, rootstock, graft, flavonoids
葡萄(Vitis vinifera L.)是世界上种植面积最大、产量最高的水果之一,种植历史悠久,约 75% 的葡萄用于酿造葡萄酒[1]。葡萄酒的风格和质量由多种物质共同决定。葡萄成熟时的类黄酮成分是决定葡萄和葡萄酒感官质量的主要因素,如颜色和苦涩味。根据结构的不同,类黄酮化合物可分为花色苷、黄酮醇和黄烷醇。花色苷是主要的呈色化合物,其含量、组成和比例以及与其他类黄酮化合物的聚合作用都会影响葡萄酒的颜色。黄酮醇作为光保护剂可保护浆果免受紫外线伤害,并可以在葡萄酒中起到辅助色素的作用[2]。黄烷醇能与花色苷形成聚合花色苷,提高葡萄酒颜色的稳定性[3]。此外,葡萄皮和种子中的黄烷醇以黄烷-3-醇和原花青素的形式存在,其中黄烷-3-醇的浓度与葡萄酒的苦味呈正相关,而原花青素的聚合度与葡萄酒的涩味密切相关[4]。因此,种植者对类黄酮物质含量适宜的葡萄需求量很大,这可以通过砧木来改变。
嫁接技术的使用始于根瘤蚜在欧洲的蔓延。葡萄和许多杂交品种需要嫁接到能抵抗根瘤蚜虫的砧木上。目前,砧木已广泛应用于葡萄栽培,它们对生物和非生物胁迫的抗性有大量文献报道[5,6]。除了这些功能外,砧木还被证明会影响葡萄树的生长和产量[7]。根据之前的一项研究,具有不同遗传背景的砧木会改变根系状态,进而影响养分吸收、树冠生物量,最后影响葡萄的成分[8]。Gutierrez-Gamboa等人[9]指出,‘美乐’嫁接到SO4上与其余七种‘美乐’嫁接葡萄相比,葡萄皮和葡萄籽中的原花青素总浓度更高,而‘美乐’/‘Gravesac’嫁接组合中的原花青素浓度较低。人们普遍认为,砧木对生殖生长和营养生长的强烈影响可能会改变丛生区的小气候和浆果成熟过程,从而改变葡萄浆果中黄酮类化合物的合成[10-12]。例如,Nelson等人[13]发现,两种嫁接葡萄的花色苷组成的差异与树冠大小的变化有关,因为树冠大小可以调节葡萄果穗接受的阳光照射。Koundouras等人[14]发现,砧木可能会通过改变果实成熟速度来改变葡萄的成分。此外,环境因素也会影响砧木的影响,如年份气候差异、土壤成分等,这些因素可能会放大或缩小砧木的变量。Han等人[15]发现,年份和砧木之间存在综合效应,而砧木在雨天和阴天较多的季节对类黄酮化合物的积累起着关键作用。之前的一些研究也表明,砧木对葡萄生长的影响与土壤水分供应、酸度等关系显著[16,17]。此外,砧木的影响也与对应的接穗品种有关。Keller等人[18]发现,3309C会诱导‘西拉’表现出低活力,反之则会诱导‘霞多丽’表现出高活力。砧木、接穗、土壤和气候条件之间存在着复杂的相互作用,因此必须针对特定地区的特定栽培品种对每种砧木进行评估。
‘小味儿多’(Vitis vinifera L. cv. Petit Verdot)原产于法国,葡萄果实富含有机酸和单宁。近年来,‘小味儿多’作为一种非常有潜力的酿酒葡萄品种在中国东部地区种植。中国东部在葡萄生长季的气候既不同于温和少雨的地中海气候,也不同于多雨寡照、炎热干燥的中国西部气候。在类似的气候条件下,关于不同砧木对‘小味儿多’葡萄类黄酮含量影响的报道很少。本研究采用HLPC-MS对中国东部地区5种砧木对‘小味儿多’葡萄果实类黄酮的影响进行了评价,旨在为实际中砧木的选择和应用提供一些参考,并为了解砧木对果实类黄酮的影响提供新的见解。
1 材料与方法
1.1 实验设计和样品采集
在中国农业大学北京上庄试验站(40°14′N, 116°20′E,海拔49米)连续进行了两年田间试验(2016-2017年)。在本研究中,‘小味儿多’(PV)于2012年分别在Paulsen 1103(1103P)、Selection Oppenheim(SO4)、Kober 5BB(5BB)、Millardet et de Grasset 101-14(101-14)和Beta上进行嫁接,株行距为2.5×1.2米,行向由南向北。试验采用随机区组设计,每个嫁接组合有三个重复,每个重复由十株葡萄藤组成。自根 PV 为对照。葡萄园叶幕形采用了改良后的VSP[19],采用短枝修剪,留梢量为每株12~15支。此外,还按照当地行业标准进行了滴灌、病虫害和营养管理。该葡萄园在葡萄生长期间的气象数据(月平均气温、日照时间和降雨量)由中国气象数据共享服务系统(http://cdc.cma.gov.cn/)提供。
2016 年和 2017 年的采收期分别为 9月25日和24日。采收时,每个重复从果穗的阳面和阴面随机采集300个浆果。采样时间为上午10:00之前,以避免中午的高温。采样后迅速测定100个浆果的理化特性,其余葡萄用于测定类黄酮化合物。
1.2 葡萄果实理化指标的测定
浆果理化指标按照中华人民共和国国家标准(GB/T15038-2006)进行测定。使用电子天平(精确度0.01 g)称量100粒浆果的重量,计算平均单粒重量。随机选取100粒浆果,用纱布包裹充分挤出果汁,用手持糖度计(PAL-1, Atago, Japan)测定其可溶性固形物含量,用pH计测定其pH值。糖积累速率(mg/berry/d)计算根据浆果的可溶性固形物含量和浆果质量计算(Antalick et al., 2021)。果汁可滴定酸用NaOH标准溶液进行滴定,结果以酒石酸计(g/L)。
1.3 葡萄果实类黄酮化合物的提取
葡萄果实类黄酮化合物的提取和检测参照之前发表的方法[20,21],果实样品经过0.22 μm的有机系膜过滤,过滤之后置于棕色液相小瓶等待上机检测。
HPLC-MS检测条件参照之前发表的方法(Wang et al., 2021):花色苷定性和定量分析使用Agilent 1200系列配备6410三重串联四级杆质谱仪(QqQ),以Poroshell 120 EC-C 18为色谱柱,柱温55°C,进样量5 μL,检测波长525 nm,流速0.4 mL/min。流动相A为0.1%(v/v)的甲酸水溶液,流动相B为含0.1%(v/v)甲酸的50/50(v/v)的甲醇乙腈溶液。洗脱程序如下:10%~100%的B相持续15 min,后运行程序5 min;离子阱质谱检测器的采集参数为电喷雾离子源,正离子模式,离子扫描范围为100~1500 m/z,雾化器压力为35 psi,干燥器流速为12 L/h,干燥气温度为350°C。非花色苷酚定性定量的HPLC-MS条件同花色苷,但采取负离子模式[78],梯度洗脱程序为0~28 min,10%~46% B;28~29 min,46%~10% B,洗脱结束后,维持10% B相冲洗5 min,平衡色谱柱。
花色苷定量以二甲花翠素-3-O-葡萄糖苷为外标物,黄酮醇以槲皮素为外标物,黄烷醇以儿茶素、表儿茶素、表棓儿茶素和表儿茶素没食子酸酯为外标物,果实中酚类化合物浓度表示为mg/kg果实鲜重(FW)。
1.4 数据处理
采用Microsoft Excel 2007进行统计分析,单因素和双因素方差分析采用SPSS 22.0,绘图采用Graphpad Prism 8.0和Simca 14.1软件。
2 结果与分析
2.1 实验地的气象特征
实验地属典型的大陆性季风气候。表 1 分析了两年的气象条件,发现5月至9月的平均气温和日照时间没有显著差异。但是,两个生长季节的降雨量差异很大。具体来说,2017 年7月至9月的降水量仅占2016年的70%,特别是2017年7月和9月的降水量不到 2016 年的四分之一。与过去10年(2008-2017年)的平均数据相比,2016年和2017年的日照时间和日均气温略有增加。在过去十年中,2016年的月平均降水量较高,这主要是由7月份的降水量较多导致的。虽然2017年的月平均降水量与过去十年一致,但8月份的降水量较高,而9月份几乎没有降水。
2.2 砧木对果实理化指标的影响
表2展示了成熟葡萄果实的重量、总可溶性固形物含量、可滴定酸和pH值。在浆果重量方面,与自根苗相比,所有嫁接葡萄树的浆果重量都略有增加。在总可溶性固形物上,砧木组合与‘小味儿多’相比没有显著影响。‘小味儿多’葡萄果汁的pH值相对于嫁接葡萄树较低,但除了PV/Beta外,两者的差异并不显著。TiA在本研究中不受砧木或季节的影响。
在本研究中,果实重量、总可溶性固形物含量和 pH值受年份影响显著。2016年浆果重量、总可溶性固形物含量和pH值较高,可能是因为2016年降水较多,日照时间较2017年短,推迟了葡萄成熟。双因素方差分析显示,只有TiA和pH显著受到了砧木和年份交互作用的影响。
验砧木与年份交互作用差异的显著性(ns 表示不显著,*p < 0.05,**p < 0.01)。
2.3 砧木对果实花色苷含量的影响
连续两年,葡萄果皮中的花色苷含量受到砧木的显著影响(图 1)。本研究表明,在两年中,‘小味儿多’自根苗的葡萄与来自PV/1103P和PV/Beta的葡萄相比,除咖啡酰化花色苷外,大多数花色苷的含量均较低。此外,PV/5BB、PV/SO4和自根苗的花色苷含量没有明显差异。而101-14砧木对花色苷含量的影响取决于不同的年份。在2016年,101-14显著降低了几种花青素的浓度,如花翠素、葡萄糖苷形式的花色苷和甲基化花色苷,但总花色苷的含量没有显著降低(图 1)。相反,与来自PV/1103P和PV/Beta的葡萄一样,嫁接到101-14 上的PV在2017年对大多数花色苷的积累有明显的积极影响,但咖啡酰化花色苷除外。除少数例外情况外,砧木对两年中不同类型花色苷比例的影响并不一致。101-14、1103P和5BB在连续两年中显著降低了葡萄糖苷形式的花色苷比例。2016年,PV嫁接砧木后,大多数葡萄乙酰化、咖啡酰化和3′-羟基化花色苷的比例较高,而香豆素化、甲氧基化和3′5′-羟基化花色苷的比例较低。2017年砧木对不同花色苷组分比例的影响与2016年相反,但甲基化花色苷除外.
2.4 砧木对葡萄黄酮醇含量的影响
砧木对葡萄果实黄酮醇含量的影响见图2。本研究发现,PV/101-14和PV/1103P组合的总黄酮醇含量显著高于自根苗,主要是由于这两种处理显著提高了杨梅酮和槲皮素的含量。此外,这两种砧木组合的葡萄果实在连续两年中大部分黄酮醇的含量都较高,只有少数例外。相反,在2016年和2017年,PV/Beta有降低黄酮醇含量的趋势,但丁香亭除外。此外,2017 年,PV/5BB和自根苗之间的黄酮醇也出现了显著差异。在两个年份中,除少数例外情况外,5BB对成熟果实中的黄酮醇含量没有显著影响。与PV/5B一样,2016年PV/SO4与自根苗相比,黄酮醇含量没有显著差异。然而,2017年,SO4砧木显著提高了大多数黄酮醇的含量,但山奈酚和西伯利亚落叶松黄酮除外。
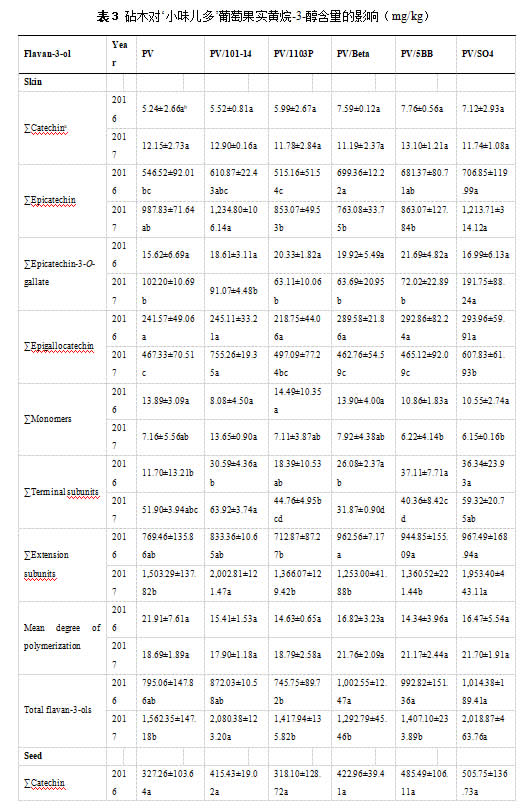
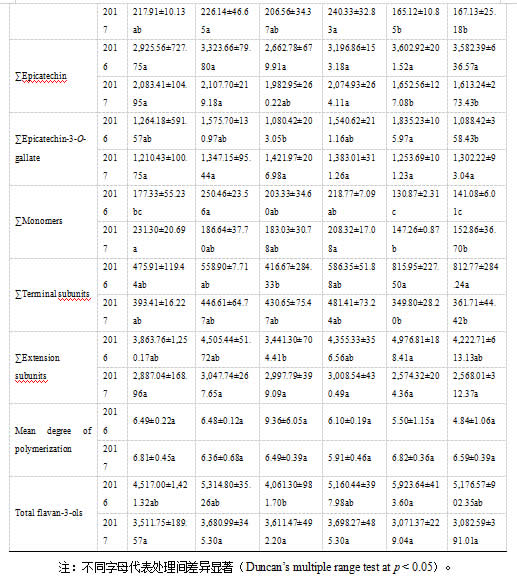
2.5 砧木对葡萄果实黄烷-3-醇含量的影响
在本研究中,砧木组合和自根苗葡萄皮中黄烷-3-醇含量存在差异(表3)。101-14和SO4仍倾向于促进葡萄皮中大部分黄烷-3-醇组分的积累,2017年的总黄烷-3-醇浓度明显高于自根苗。相反,PV/1103P对葡萄皮中黄烷-3-醇的积累有轻微的负面影响。关于葡萄籽中黄烷-3-醇的含量,砧木组合和自根苗之间没有显著差异(表 3)。其中,1103P对连续两季的黄烷-3-醇含量几乎没有影响。此外,101-14和Beta对本研究中葡萄籽中黄烷-3-醇的含量也有轻微的积极影响。在果皮和种子的平均聚合度方面,自根苗和各砧木组合在两个连续年份中没有发现显著差异。
2.6 葡萄果实类黄酮化合物的主成分分析
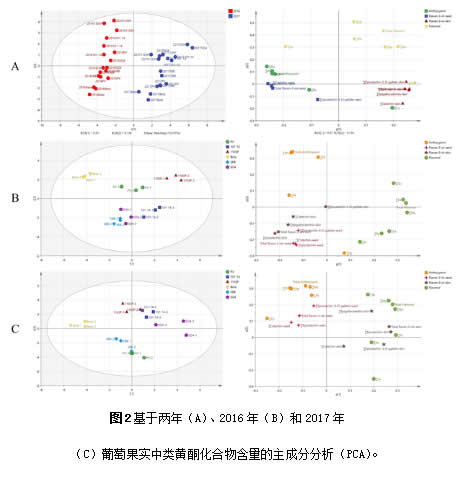
如图 3A 所示,主成分分析(PCA)是进一步明确‘小味儿多’砧木组合和自根苗成熟葡萄中类黄酮化合物特征的重要方法。值得注意的是,在PCA模型中,年份是区分样品的主要因素,而砧木组合和自根苗并没有明显的区分。前两个主成分分别解释了总方差的 53.0% 和 15.4%。第一个主成分(PC1)将2016年和2017年两个年份区分开。载荷图显示,2016年的葡萄果实中花色苷的含量更高,种子中黄烷-3-醇的含量更高,而2017年的葡萄皮中黄酮醇和黄烷-3-醇的浓度更高。
为了减少年份的影响,2016年和 2017年分别根据类黄酮化合物的原始浓度进行了 PCA,以研究不同砧木的生理特性(图 3)。对于2016年的葡萄,PC1占总方差的31.2%。PV/1103P和PV/101-14组合的样品位于PC1的正半轴部分,而PV/Beta组合的样品位于PC1的负半轴部分。黄酮醇对PC1有正向影响,而花色苷和黄烷-3-醇对PC1有负面影响。PC2解释了总方差的19.4%,将PV/Beta和PV/1103P分为一组,将其他砧木分为另一组,PV位于两组之间。PV/1103P和PV/Beta的葡萄含有更多的花色苷,而其他砧木组合的葡萄则含有更多的花翠素、异鼠李素、丁香亭和西伯利亚落叶松黄酮以及更多的黄烷-3-醇。2017年,PC1 解释了32.9%的变异,并将样品分为三组。PV/SOPV/101-1组合的葡萄果皮中含有更多的黄酮醇和黄烷-3-醇,PV/Bet组合的葡萄种子中含有更多的黄烷-3-醇和花青素。PC2占总变异的26.1%,区分了四种砧木组合,如PV/1103P与PV和PV/5BB。结合两年来的一致结果,这些结果表明1103P促进了光伏浆果果皮中花色苷和黄酮醇的积累。Beta有利于‘小味儿多’葡萄果皮中花花色苷和种子中黄烷-3-醇的生物合成,不利于果皮中黄酮醇的生物合成。101-14能够提高‘小味儿多’葡萄果皮中黄酮醇和黄烷-3-醇的浓度。SO4增加了黄烷醇在‘小味儿多’葡萄果皮中的积累。
3 讨论
这项连续两年的田间试验研究了五种砧木与自根葡萄树相比对‘小味儿多’葡萄理化指标和类黄酮的影响。
采收期浆果成分是葡萄和葡萄酒品质的主要决定因素,取决于各自的环境、砧木和栽培之间复杂的相互作用[9,24-26]。在气候变暖的背景下,葡萄树需要应对不断增加的水分胁迫,因此砧木和水分状况在其他因素中显得更为重要[27]。在本研究中,2016年和2017年的降雨量表现出很大差异。2016年7月至9月的降水量明显高于2017年,尤其是2017年7月和9月的降水量不到2016年的四分之一。值得注意的是,葡萄的成熟期从7月开始,采收期从9月开始,这段时间降雨对葡萄的成熟和成分有很大影响。因此,与2017年相比,2016年降水较多,日照时间较短,导致葡萄成熟度适中。物候期推迟可能是导致2016年浆果重量、TSS、pH值升高和TiA降低的原因。
一般来说,砧木会通过改变葡萄树的营养和生殖生长来影响葡萄的成熟速度。如之前的研究,‘赤霞珠’嫁接到1103P上时,其生营养生长能力较强,导致‘赤霞珠’葡萄延迟成熟[15,28]。不过,同一砧木对不同接穗栽培品种的影响可能不同[29]。从葡萄的理化参数来看,砧木对‘小味儿多’的浆果成熟速度没有影响。有研究报告称,砧木对果实理化指标的影响较小,这与我们的研究结果一致[30]。此外,最近也有关于砧木对成熟葡萄理化指标的影响的报道,但这些影响并未持续改变浆果重量、TSS、pH值和TiA [9,15,24]。值得注意的是,砧木对接穗葡萄pH值和TiA的影响与钾的吸收有关,因为钾会沉淀果汁中的酒石酸盐,导致酒石酸减少[31]。然而,葡萄对钾的吸收能力受到接穗栽培品种、砧木基因型和土壤条件相互作用的影响,土壤中钾的供应量越大,就越能掩盖不同砧木对钾吸收能力的差异[16]。在本研究中,嫁接葡萄的TiA和果汁的pH值没有显著差异,这可能与我们之前报道的葡萄园土壤中富含大量的钾有关[30]。
花色苷分布在红葡萄的果皮中,是红葡萄和葡萄酒着色的关键物质[32]。与之前的研究一致[33],‘小味儿多’中的乙酰化和甲基化花色苷是葡萄花色苷的主要成分。然而,在‘佳丽酿’和‘歌海娜’等几个品种中,花色苷总量的主要成分是香豆酰化形式[34,35]。这表明不同栽培品种的花色苷含量存在一些差异。有趣的是,在酰化花色苷中,咖啡酰化花色苷的浓度和比例在很大程度上受砧木的影响,这与最近的一项研究一致[30]。虽然没有找到可能的原因,但我们的发现是咖啡酰化花色苷受生物调节的新证据。在本研究确定的砧木中,1103P和Beta有利于葡萄皮中花色苷浓度的积累。然而,葡萄栽培文献通常将1103P归类为生命力极强的砧木,这种砧木往往会增加叶果比,削弱花色苷的生物合成[15,28,36]。这种不一致的结果主要是由不同的接穗品种和环境因素造成的。不同的接穗品种对砧木的亲和力不同,对复杂环境的反应也不同,这些都会影响花色苷的合成。Beta是华东地区常用的砧木,因为它具有很强的抗寒性[37]。有报道称,将Beta砧木嫁接到‘夏黑’上会增加花色苷浓度,这与本研究一致[38]。除少数情况外,101-14对花色苷浓度的影响主要受季节因素的影响。此外,5BB和SO4对花色苷浓度的影响不大,这与之前的研究一致[14,30]。值得注意的是,本研究发现,在降水较少、光照强度较大的生长季节,砧木组合的花色苷含量更为明显。最近的一项研究[30]对赤霞珠也有类似发现。这是因为强光会上调花色苷生物合成途径关键基因的表达。
黄酮醇是重要的辅色素,可以稳定葡萄酒的颜色。在我们的研究中,5BB对成熟浆果中的黄酮醇浓度没有显著影响,与之前的研究一致[30]。此外,在两个生长季节,PV/101-14和PV/1103P组合果皮中的黄酮醇浓度较高。槲皮素是‘小味儿多’葡萄中含量最高的黄酮醇,101-14和1103P在显著增加了槲皮素的含量。这些结果表明,嫁接到‘小味儿多’上的101-14和1103P有促进黄酮醇生物合成的趋势。值得注意的是,黄酮醇对光特别敏感,在葡萄皮中具有紫外线保护作用[26]。一项研究发现,1103P增加了修剪重量,导致光照不足,从而降低了‘赤霞珠’中黄酮醇的浓度[15]。此外,在这项研究中,Beta还大大降低了果皮中黄酮醇的浓度,而一项研究报告称,这种砧木会导致接穗活力低下,从而增加浆果的受光度[37]。
大部分黄烷-3-醇积累在葡萄浆果的种子和果皮中。根据以往的研究,砧木对黄烷-3-醇的浓度没有显著影响,这表明黄烷-3-醇的浓度相对稳定,不易受栽培方法的影响[15,39,40]。然而,在本研究中,嫁接到101-14上的小味而多葡萄在两个年份中黄烷-3-醇的浓度都有一定程度的增加,这与之前的报告一致[9]。此外,年份仍然是造成嫁接葡萄和自根葡萄中黄烷-3-醇浓度差异的主要因素。在本研究中,2017年光照充足、雨水较少,葡萄中的黄烷-3-醇浓度较低,而2016年光照较少、雨水较多,葡萄中的黄烷-3-醇浓度较高。这种差异主要是由黄烷-3-醇的特殊积累模式造成的。最近的一项研究表明,葡萄果实中的黄烷-3-醇通常在葡萄成熟期达到最高浓度,然后呈下降趋势[41]。生长季光照充足,雨水较少,加快了黄烷-3-醇的分解速度,进而导致该年份成熟浆果中的黄烷-3-醇浓度较低[42]。此外,由于葡萄酒的涩味不仅取决于原花青素的浓度,还取决于其结构,因此平均聚合度(mDP)也是一个经常被关注的指标。与之前的报告一致,果皮原花青素的mDP要高于种子原花青素[43]。Gutiérrez-Gamboa等人[35] 发现,101-14和SO4增加了‘美乐’葡萄果皮原花青素的mDP。但是,这一结果在‘小味儿多’中并没有得到明确的证实。
4 结论
近年来,‘小味儿多’葡萄在中国东部的一些产区得到了试验和推广,但关于砧木对‘小味儿多’葡萄类黄酮化合物的影响的报道却很有限。一般来说,不同的砧木可以延迟或加速葡萄果实的成熟,但在本研究中,‘小味儿多’葡萄的砧木并没有明确的证据。年份是影响浆果理化指标的决定性因素,而砧木对浆果理化指标没有显著影响,只有Beta能显著提高pH值。砧木对‘小味儿多’葡萄中类黄酮化合物的积累起了重要作用,尤其是花色苷和黄酮醇。1103P在两季中对浆果皮中花色苷和黄酮醇的积累起到了积极作用。在这项研究中,Beta能明显增加浆果果皮中的花色苷浓度,同时也对除丁香亭以外的黄酮醇的生物合成产生不利影响。嫁接到101-14砧木上的葡萄,果皮中黄酮醇的浓度较高,但仍低于PV/1103P组合。葡萄中黄烷-3-醇的含量相对稳定,只有101-14在两年中黄烷-3-醇的含量略有增加。此外,不同年份的气候差异可能会掩盖某些砧木栽培品种的影响。SO4对葡萄皮中花色苷和黄酮醇的影响易受年份因素的影响,在连续两年中的结果并不一致。PCA结果表明,PV/5BB的类黄酮化合物含量与‘小味儿多’自根苗最接近,其余砧木品种与自根苗存在差异。总之,本研究为中国东部地区‘小味儿多’的嫁接栽培提供了理论依据。